Precieze gene bewerken vormt de hoeksteen van grote biomedische ontwikkelingen die betrekking hebben op de ontwikkeling van nieuwe strategieën om genetische ziekten te bestrijden. Op de voorhoede van gen-bewerken technologieën is de methode afhankelijk van het gebruik van de clustered regularly –Iknterspaced short palindromic repeats (CRISPR) / Cas9-systeem dat aanvankelijk werd geïdentificeerd Als een onderdeel van bacteriële immuniteit tegen de invasie van virale genetische materiaal (herzien in verwijzingen1,2). Een groot voordeel van het systeem van CRISPR/Cas9 over andere gen-bewerkingsgereedschap, zoals zink-vinger nucleasen (ZFNs) en transcriptie activator-achtige effector nucleasen (TALENs) (herzien in verwijzing3), is de relatieve eenvoud van plasmide ontwerp en bouw van de CRISPR componenten — een functie die de uitbreiding van het gen-bewerkingen van enkele gespecialiseerde laboratoria tot een veel ruimere onderzoeksgemeenschap heeft aangedreven. Bovendien hebben de eenvoud van CRISPR/Cas9 programmering en haar capaciteit voor multiplexed doel erkenning voor het verder aangewakkerd zijn populariteit als een rendabele en easy-to-use technologie. Onder de verschillende methoden beschikbaar voor onderzoekers te leveren van dergelijke componenten bewerken van gen naar cellen, blijven virale vectoren verreweg het populairste en meest efficiënte systeem.
Lentivirale vectoren (LVs) opgedoken als het voertuig van keuze te leveren van de onderdelen van CRISPR/Cas9 systeem in vivo voor uiteenlopende toepassingen4,5,6,7. Aantal belangrijke functies maken LVs een populaire keuze voor dit proces met inbegrip van hun vermogen om te verdelen en niet-delende cellen, lage immunogeniciteit zowel minimale cellulaire toxiciteit (herzien in verwijzing8) besmetten. Dientengevolge, is LV-gemedieerde gentherapie werkzaam in de behandeling van infectieziekten, zoals HIV-1, HBV en HSV-1, alsook in de correctie van fouten ten grondslag liggen aan erfelijke ziekten bij de mens, zoals mucoviscidose en neo-vasculaire macula degeneratie 4 , 5 , 7 , 9 , 10 , 11. Bovendien, LVs effectief zijn gewijzigd om een multiplex gene bewerkingsfuncties op verschillende genomic loci met behulp van een enkele vector systeem12.
Echter de inherente eigenschap van LVs te integreren in het genoom van de gastheer kan mutagene en handicaps vaak hun nut als transgenic leverende voertuigen, met name in klinische instellingen. Bovendien, aangezien stabiel geïntegreerde LVs hun transgenen op een duurzaam hoog niveau uiten, dit systeem is niet geschikt voor de levering van gen-bewerken onderdelen zoals CRISPR/Cas9; overexpressie van Cas9-gids RNA (gRNA), en soortgelijke eiwitten zoals ZFNs, worden geassocieerd met verhoogde niveaus van off-target effecten, waaronder ongewenste mutaties13,14,15,16 , 17 en het potentieel van de cytotoxiciteit18kan verbeteren. Daarom, om precieze gen-bewerken met minimale uit-target effecten, is het noodzakelijk om ontwerpsystemen waarmee voor de voorbijgaande expressie van gen editing-onderdelen.
In de afgelopen jaren een scala aan levering platforms zijn ontwikkeld om te Transient express CRISPR/Cas9 in cellen16,19,20,21 (herzien in verwijzing22). Het gaat hierbij om methoden die afhankelijk zijn van direct invoering van gezuiverde Cas9 samen met de juiste gids RNAs in cellen, die werd getoond om meer effectief bij gerichte gen-bewerken in vergelijking met de plasmide gemedieerde transfectie16. Studies hebben aangetoond dat ribonucleoprotein (RNP) complexen bestaande uit begeleiden RNA/Cas9 deeltjes zijn snel overgedragen na bemiddeling van DNA decollete op hun doelen, suggereert dat op korte termijn uitdrukking van deze componenten is voldoende om te bereiken robuuste gene16bewerken. Denkbaar, niet-integratie van virale vector platforms zoals adeno-geassocieerde virale vectoren (AAVs) bieden een levensvatbaar alternatief voor het bewerken van gen machines leveren aan cellen. Helaas, AAV capsids aanzienlijk lager verpakking mogelijkheid dan LVs bezitten (< 5kb), die ernstig belemmert hun vermogen voor het inpakken van de multi-component CRISPR toolkit binnen een enkele vector (herzien in verwijzing8). Het is vermeldenswaard dat toevoeging van verbindingen die remmen Histon deacetylases (b.v., natrium butyraat23) of belemmeren van de celcyclus (b.v., cafeïne24) is aangetoond dat het verhogen van lentivirale titers. Ondanks de recente vooruitgang, zijn de Transiënte expressiesystemen ontwikkeld tot nu toe nog steeds belemmerd door verschillende tekortkomingen, zoals lagere productie-efficiëntie, die tot verminderde virale titers, en lage transductie efficiëntie van de virussen die zijn gegenereerd leiden door dergelijke benaderingen25.
Integrase-deficiënte lentivirale vectoren (IDLVs) vormen een grote vooruitgang in de ontwikkeling van gen-leverende voertuigen, zoals zij het vermogen van de verpakking van LVs met het extra voordeel van AAV-achtige episomal onderhoud in cellen combineren. Deze voorzieningen helpen IDLVs grotendeels het omzeilen van de belangrijke kwesties in verband met het integreren van vectoren, vis-à-vis continu overexpressie van potentieel genotoxisch elementen en integratie-gemedieerde mutageniteit. Het werd eerder aangetoond dat IDLVs met succes kan worden aangepast voor het verbeteren van de episomal gen expressie26,27. Met betrekking tot de CRISPR/Cas9 IDLV-gemedieerde levering beperkt lage productie titers en lagere expressie van isomer overgedragen genomen ten opzichte van integrase-bedreven lentivirale systemen hun nut als bona fide tools voor het leveren van genoom-bewerken transgene constructies. We onlangs aangetoond dat zowel transgenic expressie en virale titers IDLV productie gekoppeld aanzienlijk worden verhoogd door de opneming van bindende sites voor de transcriptiefactor Sp1 binnen de virale expressie cassette28. De gewijzigde IDLVs ondersteund krachtig CRISPR-gemedieerde gene zowel in vitro (HEK-293T cellen) en in-vivo (in post mitotische hersenen neuronen), terwijl het minimale uit-target mutaties ten opzichte van de bijbehorende ICLV-gemedieerde inducerende bewerken systemen28. Globaal, ontwikkelden we een roman, compact, alles-in-één CRISPR toolkit uitgevoerd op een platform van IDLV en geschetst van de verschillende voordelen van het gebruik van dergelijke een leveringsvoertuig voor het bewerken van de verbeterde gen.
Hier, is het protocol van de productie van de IDLV-CRISPR/Cas9-systeem beschreven, met inbegrip van de verschillende stappen in de vergadering, zuivering, concentratie en titratie van IDLVs, evenals strategieën voor het valideren van het bewerken van gene werkzaamheid van deze vectoren. Dit protocol is eenvoudig schaalbaar naar de behoeften van verschillende onderzoekers en is ontworpen om met succes tot LV vectoren met titers in het bereik van 1 x 1010 eenheden (TU) transducing / mL. De vectoren gegenereerd door dit protocol kunnen worden gebruikt om verscheidene verschillende soorten cellen, met inbegrip van moeilijk-aan-transduce embryonale stamcellen, hematopoietische cellen (T-cellen en macrofagen), en gekweekt en in vivo– efficiënt te besmetten ingespoten neuronen. Bovendien is het protocol is even geschikt voor de productie van integrase-bevoegde lentivirale vectoren in vergelijkbare hoeveelheden.
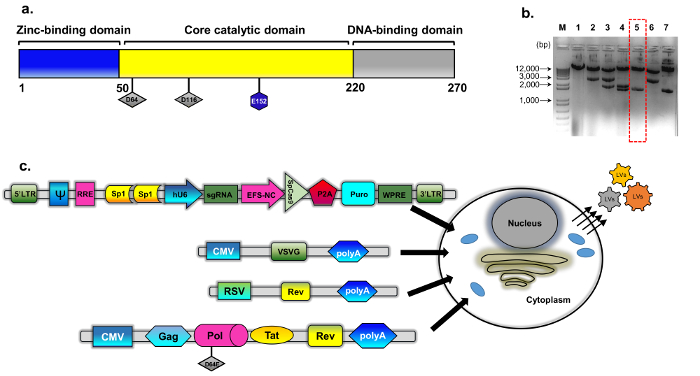
Figuur 1: IDLV verpakking. (a) schematische voorstelling van de wild type integrase eiwit (b) de gemodificeerde plasmide is afgeleid van psPAX2 (Zie methoden, plasmide constructie voor details). Vertegenwoordiger agarose gel afbeelding van klonen gescreend voor gemuteerde integrase klonen. Bereid met behulp van een standaard plasmide DNA isolatie Mini-Kit DNA-monsters werden geanalyseerd door de spijsvertering met EcoRV en SphI. De kloon van goed verteerd (nummer 5, onderbroken rood kader) werd verder gecontroleerd door directe (Sanger) volgorde van de vervanging van de D64E in INT. De integrase-deficiënte verpakking cassette heette pBK43. (c) schematische voorstelling van het protocol van de voorbijgaande transfectie werkzaam voor het genereren van IDLV-CRISPR/Cas9 vectoren, toont 293T cellen transfected met VSV-G, verpakking en transgenic cassettes (plasmide van de alles-in-één van de Sp1-CRISPR/Cas9). Virale deeltjes die bud uit van de celmembraan bevatten de full-length RNA van de vector (uitgedrukt uit de transgenic cassette). De tweede generatie van het systeem van de IDLV-verpakking werd gebruikt, waaronder de regelgevende eiwitten Tat en Rev. Rev expressie is verder aangevuld van een aparte cassette (RSV-REV-plasmide). Abbrev: LTR-lange-terminal herhalen, VSV-G, vesiculaire stomatitis virus G-eiwit, pCMV-cytomegalovirus promotor; Rous Sarcoom virus (RSV) promotor; RRE-(Rev respons-Element). Andere regulerende elementen op de expressie cassette opnemen Sp1-bandplaatsen, Rev Response element (RRE), Woodchuck Hepatitis Virus Posttranscriptional regelgevende Element (WPRE), een kern-rek factor 1α promotor (EFS-NC), de vector-verpakking element ψ (psi), menselijke Cytomegalovirus (hCMV) promotor en menselijke U6 promotor. Klik hier voor een grotere versie van dit cijfer.
