Gen präzise Bearbeitung bildet den Grundstein biomedizinische Fortschritte, die die Entwicklung neuer Strategien zur Bekämpfung genetischer Krankheiten betreffen. An die Spitze der gen-Bearbeitungs-Technologien ist die Methode unter Berufung auf die Nutzung der clustered rregelmässig –IchNterspaced sHort pAlindromic REpeats (CRISPR) / Cas9-System, das zunächst identifiziert wurde als Bestandteil des bakteriellen Immunität gegen die Invasion der viralen Erbsubstanz (rezensiert in Referenzen1,2). Ein wesentlicher Vorteil des CRISPR/Cas9-Systems gegenüber anderen gen-editing-Tools, wie z. B. Zinkfinger-Nukleasen (ZFN) und Transkription Aktivator-ähnliche Effektor Nukleasen (TALENs) (rezensiert in Referenz3), ist die relative Einfachheit der Plasmid-Design und Bau von CRISPR-Komponenten – ein Feature, das den Ausbau der gen-Bearbeitung von wenigen Speziallabors für eine viel breitere Forschungsgemeinschaft angetrieben hat. Darüber hinaus haben die Einfachheit der CRISPR/Cas9 Programmierung und seine Kapazität für Multiplex Zielerfassung seine Popularität als eine kostengünstige und einfach zu bedienende Technologie weiter angeheizt. Unter den verschiedenen Methoden zur Verfügung, um Forscher solche gen-Bearbeiten von Komponenten an Zellen zu liefern bleiben virale Vektoren bei weitem die beliebteste und effizientes System.
Lentivirale Vektoren (LVs) entstanden als das Fahrzeug der Wahl für die Komponenten des CRISPR/Cas9 System in Vivo für unterschiedlichste Anwendungen4,5,6,7liefert. Mehrere wichtige Funktionen machen LVs eine beliebte Wahl für diesen Prozess, einschließlich ihrer Fähigkeit, sowohl trennende und nicht teilenden Zellen, niedrige Immunogenität und minimale zelluläre Toxizität (rezensiert in Referenz8) zu infizieren. Infolgedessen wurde LV-vermittelte Gentherapie in der Behandlung von Infektionskrankheiten wie HIV-1, HBV und HSV-1, sowie bei der Korrektur von Mängeln, die zugrunde liegenden menschlichen Erbkrankheiten, wie Mukoviszidose und Neo-Kreislauf Makuladegeneration eingesetzt 4 , 5 , 7 , 9 , 10 , 11. Außerdem, LVs wurden effektiv geändert um Multiplex-gen an unterschiedliche genomic Loci mit einem einzigen Vektor System12Bearbeitung ausführen.
Jedoch kann die inhärente Eigenschaft des LVs in das Wirtsgenom integrieren mutagen und oft behindert ihre Nützlichkeit als Transgen Lieferfahrzeuge, vor allem im klinischen Umfeld. Da zudem stabil integriert LVs ihre transgene auf einem nachhaltig hohen Niveau zum Ausdruck bringen, ist dieses System ungeeignet für die Lieferung von gen-Bearbeiten von Komponenten wie CRISPR/Cas9; Überexpression des Cas9-Guide RNA (gRNA) und ähnliche Proteine wie ZFN, sind verbunden mit erhöhten Konzentrationen von Ziel-Effekte, darunter unerwünschte Mutationen13,14,15,16 , 17 und können potenziell Zytotoxizität18verbessern. Daher unbedingt um präzise zu erreichen gen-Bearbeitung mit minimal Ziel Effekte, Design-Systemen, die für die transiente Expression des Gens, die Bearbeitung von Komponenten ermöglichen.
In den letzten Jahren wurden eine Vielzahl von Plattformen entwickelt, um vorübergehend CRISPR/Cas9 in Zellen16,19,20,21 (rezensiert in Referenz22) zum Ausdruck bringen. Dazu gehören Methoden, die direkt Einführung von gereinigten Cas9 zusammen mit der passenden Guide-RNAs in Zellen, die gezeigt wurde, effektiver auf gezielte gen-Bearbeitung im Vergleich zu Plasmid-vermittelte Transfektion16abhängig. Studien haben gezeigt, dass Ribonucleoprotein (RNP) komplexe, bestehend aus Leitfaden RNA/Cas9 Partikel werden schnell umgedreht nach Vermittlung Spaltung der DNA auf ihre Ziele, was darauf hindeutet, dass kurzfristige Ausdruck dieser Komponenten zu erzielen ist robuste gen16bearbeiten. Denkbar, nicht-Integration von viralen Vektoren Plattformen wie Adeno-assoziierten viralen Vektoren (Flugabwehrpanzer) bieten eine echte Alternative um gen-Bearbeitung Maschinen an Zellen zu liefern. Leider AAV Capsids besitzen deutlich geringere Verpackung Fähigkeit als LVs (< 5kb), die schwer behindert ihre Fähigkeit, das Mehrkomponenten CRISPR-Toolkit innerhalb einer einzigen Vektor (rezensiert in Referenz8) zu verpacken. Es ist erwähnenswert, dass die Zugabe von Verbindungen, die Histon Deacetylases (z.B. Natrium Butyrat23) hemmen oder verhindern den Zellzyklus (z.B. Koffein24) gezeigt worden, um Lentivirale Titer zu erhöhen. Trotz der jüngsten Fortschritte sind die transiente Expression-Systeme so weit entwickelt noch mehrere Mängel auf, z. B. geringere Produktionseffizienz, behindert durch führen zu reduzierten virale Titer und niedrigen Transduktion Effizienz erzeugt durch Viren solche Ansätze25.
Integrase-defizienten Lentivirale Vektoren (IDLVs) stellen einen großen Fortschritt in der Entwicklung von gen-Lieferfahrzeuge, wie sie die Verpackung-Fähigkeit des LVs mit dem zusätzlichen Vorteil der AAV-artigen episomal Wartung in Zellen verbinden. Diese Funktionen helfen IDLVs, die die wichtigsten Probleme im Zusammenhang mit Integration von Vektoren, Vis À Vis kontinuierliche Überexpression von potenziell genotoxische Elementen und Integration-vermittelten Mutagenität weitgehend zu umgehen. Es wurde bereits gezeigt, dass IDLVs erfolgreich zur Verbesserung der episomal gen Ausdruck26,27geändert werden kann. In Bezug auf IDLV-vermittelten CRISPR/Cas9 Lieferung beschränkt niedrige Titer und niedriger Ausdruck von Episome getragen Genomen relativ Integrase-kompetente Lentivirale Systeme ihre Nützlichkeit als Bona Fide -Tools für die Bereitstellung von Genom-Bearbeitung transgene Konstrukte. Wir zeigten kürzlich, dass Transgene Ausdruck und virale Titer IDLV Produktion durch die Einbeziehung von Bindungsstellen für den Transkriptionsfaktor Sp1 in der viralen Ausdruck Kassette28erheblich verbessert werden. Die modifizierte IDLVs unterstützt robust CRISPR-vermittelten gen Bearbeitung sowohl in Vitro (in HEK-293T Zellen) und in Vivo (im Post-mitotische Gehirnneuronen), während minimalste Ziel Mutationen im Vergleich zu den entsprechenden ICLV-vermittelte Induktion Systeme-28. Insgesamt entwickelten wir eine neuartige, kompakte, all-in-One CRISPR-Toolkit auf einer IDLV Plattform durchgeführt und erläutert die verschiedenen Vorteile der Verwendung solch ein Transportvehikel für verbesserte gen zu bearbeiten.
Hier ist das Produktionsprotokoll des IDLV-CRISPR/Cas9-Systems beschrieben, darunter die verschiedenen Schritte in der Montage, Reinigung, Konzentration und Titration von IDLVs sowie Strategien, die gen-Bearbeitung Wirksamkeit dieser Vektoren zu validieren. Dieses Protokoll ist leicht skalierbar, um die Bedürfnisse der verschiedenen Forschern und wurde entwickelt, um erfolgreich LV Vektoren mit Titer im Bereich von 1 x 1010 transducing Einheiten (TU) erzeugen / mL. Die Vektoren generiert durch dieses Protokoll können genutzt werden, um effizient mehrere verschiedene Zelltypen, einschließlich schwer transduzieren embryonale Stammzellen, blutbildenden Zellen (T-Zellen und Makrophagen) und kultiviert und in Vivozu infizieren- eingespritzte Neuronen. Darüber hinaus ist das Protokoll für die Produktion der Integrase-kompetente Lentivirale Vektoren in ähnlichen Mengen gleich gut geeignet.
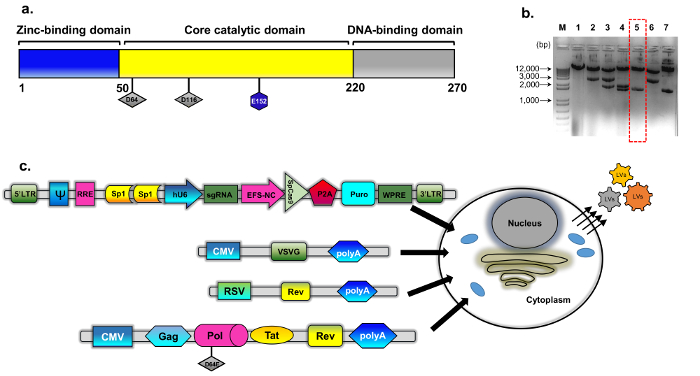
Abbildung 1: IDLV Verpackung. (a) schematische Darstellung der Wildtyp Integrase Protein (b) das modifizierte Plasmid wurde von psPAX2 abgeleitet (siehe Methoden, Plasmid-Konstruktion für Details). Vertreter Agarose Gelbild geschirmt für mutierte Integrase Klone Klone. DNA-Proben mit den standard Plasmid DNA-Isolierung Mini-Kit zubereitet wurden durch Vergärung mit EcoRV und SphI analysiert. Der richtig verdaut Klon (Nummer 5, gestrichelten roten Feld) wurde weiter durch Sequenzierung der direkten (Sanger) für die D64E Substitution in INTüberprüft. Die Integrase-defizienten Verpackung Kassette wurde pBK43 genannt. (c) schematische Darstellung der transiente Transfektion Protokoll eingesetzt, um generieren IDLV-CRISPR/Cas9 Vektoren zeigen, dass 293T Zellen mit VSV-G, Verpackungen und Transgen Kassetten (Sp1-CRISPR/Cas9 all-in-One Plasmid) transfiziert. Viruspartikel, die heraus von der Zellmembran Knospe enthalten die Full-length RNA des Vektors (ausgedrückt aus der Transgen-Kassette). Die zweite Generation der IDLV-Verpackungssystem diente, worunter die regulatorische Proteine Tat und Rev Rev Ausdruck wird von einer separaten Kassette (RSV-REV-Plasmid) ergänzt. Abbrev: LTR Long Terminal repeat, VSV-G, Stomatitis vesicularis Virus G-Protein, pCMV-Zytomegalievirus Veranstalter; Rous Sarkom-Virus (RSV) Veranstalter; RRE-(Rev-Response-Element). Weitere regulatorische Elemente auf der Expressionskassette gehören Sp1-Bindungsstellen, Rev Response-Element (RRE), Murmeltier Hepatitis Virus posttranskritionelle regulatorische Element (WPRE), Kern-Dehnung Faktor 1α Förderer (EFS-NC), der Vektor-Verpackung Element-ψ (Psi), menschliches Cytomegalovirus (hCMV) Veranstalter und menschlichen U6 Förderer. Bitte klicken Sie hier für eine größere Version dieser Figur.
