Edição de gene preciso constitui a pedra angular das grandes avanços biomédicos que envolvem o desenvolvimento de novas estratégias para combater doenças genéticas. Na vanguarda das tecnologias de edição de gene é o método baseando-se no uso do cabrilhantado regularly –eunterspaced short palindromic repeats (CRISPR) / sistema de Cas9 que foi inicialmente identificado como um componente da imunidade bacteriana contra a invasão de material genético viral (revisto em referências1,2). Uma grande vantagem do sistema CRISPR/Cas9 sobre outras ferramentas de edição de gene, como nucleases dedo de zinco (ZFNs) e as nucleases efetoras como ativador da transcrição (TALENs) (revistas em referência3), é a relativa simplicidade de design do plasmídeo e construção de componentes CRISPR — um recurso que tem alimentado a expansão do gene-edição de alguns laboratórios especializados para uma comunidade de pesquisa muito mais ampla. Além disso, a simplicidade de programação CRISPR/Cas9 e sua capacidade de reconhecimento do alvo multiplexado alimentaram ainda mais sua popularidade como uma tecnologia de baixo custo e fácil de usar. Entre os vários métodos disponíveis para pesquisadores entregar tais componentes de edição de gene para as células, vetores virais continuam a ser de longe o sistema mais popular e eficiente.
Vetores de Lentivirus (LVs) têm emergido como o veículo de escolha para entregar os componentes do CRISPR/Cas9 sistema na vivo para diversas aplicações4,5,6,7. Vários recursos chaves fazem LVs uma escolha popular para este processo, incluindo a sua capacidade de infectar células de divisão e não dividindo, baixa imunogenicidade e mínima toxicidade celular (revisto em referência8). Como resultado, a terapia gênica mediada por LV tem sido empregada em tratamentos de doenças infecciosas, tais como o HBV, HIV-1 e HSV-1, bem como na correção de defeitos subjacentes humanas doenças hereditárias, como fibrose cística e degeneração macular neo-vascular 4 , 5 , 7 , 9 , 10 , 11. Além disso, a LVs foram efetivamente modificados para realizar a edição de gene multiplex em distinta loci genômicos usando um vetor único sistema12.
No entanto, a propriedade inerente de LVs para integrar o genoma do hospedeiro pode ser mutagênico e frequentemente desfavorecem sua utilidade como veículos de entrega do transgene, especialmente em situações clínicas. Além disso, desde que integrado estàvel LVs expressam seus transgenes níveis elevados sustentavelmente, este sistema é suficientemente adaptados para a entrega de componentes de edição de gene como CRISPR/Cas9; superexpressão de RNA Cas9-guia (gRNA) e proteínas similares, tais como ZFNs, estão associados com níveis elevados de efeitos fora do alvo, que incluem mutações indesejáveis13,14,15,16 , 17 e que potencialmente pode melhorar a citotoxicidade18. Portanto, para obter precisão edição de gene com efeitos mínimos fora do alvo, é imperativo para design de sistemas que permitem a expressão transiente de gene editando componentes.
Nos últimos anos, uma variedade de plataformas de entrega foram desenvolvidos para expressar transitoriamente CRISPR/Cas9 em células16,19,20,21 (revisto em referência22). Estes incluem métodos que dependem diretamente introduzir Cas9 purificado, juntamente com as guia apropriada RNAs em células, que foi mostrado para ser mais eficaz no gene alvo-edição em comparação com a transfeccao mediada por plasmídeo16. Estudos têm demonstrado que ribonucleoprotein (RNP) complexos consistindo de guia RNA/Cas9 partículas são rapidamente virou depois da mediação de clivagem de DNA em seus alvos, sugerindo que a expressão a curto prazo desses componentes é suficiente para alcançar gene robusto edição16. É concebível, não-integração de plataformas de vetor viral como vetores de vírus adeno-associado (AAVs) podem fornecer uma alternativa viável para entregar máquinas edição de gene em células. Infelizmente, capsids AAV possuem capacidade de embalagem significativamente menor do que o LVs (< 5KB), que dificulta severamente sua capacidade para empacotar o toolkit CRISPR multi-componentes dentro de um único vetor (revisto em referência8). É interessante notar que a adição de compostos que inibem a histona deacetilases (por exemplo, o butirato de sódio23) ou impedir o ciclo celular (por exemplo, cafeína24) foram mostrados para aumentar a concentração de particulas de Lentivirus. Apesar dos progressos recentes, os sistemas de expressão transiente desenvolvidos até agora ainda são impedidos por várias deficiências, tais como a baixa eficiência da produção, que levam a reduzida concentração viral e transdução de baixa eficiência dos vírus gerados através de tais abordagens25.
Vetores de Lentivirus integrase deficiente (IDLVs) representam um grande avanço no desenvolvimento de veículos de entrega de gene, como eles combinam a capacidade de empacotamento de VLS com o benefício adicionado de AAV-como manutenção epissomal nas células. Esses recursos ajudam IDLVs largamente contornar as principais questões associadas com a integração de vetores, vis-à-vis a superexpressão contínua do potencial genotóxico de elementos e mutagenicidade mediada por integração. Foi previamente demonstrado que IDLVs podem ser modificadas com êxito para melhorar a expressão de gene epissomal26,27. Com relação à entrega de mediada por IDLV CRISPR/Cas9, títulos de baixa produção e menor expressão dos genomas episome-suportado em relação a sistemas de Lentivirus integrase-proficiente limita sua utilidade como ferramentas de bona fide para a entrega de genoma-edição construções de transgénicas. Recentemente demonstramos que tanto a expressão do transgene e títulos virais associados à produção de IDLV são significativamente melhorados através da inclusão de binding sites para o fator de transcrição Sp1 dentro da gaveta de expressão viral28. Os IDLVs modificados robustamente suportado gene mediada por CRISPR edição tanto in vitro (em células HEK-293T) e na vivo (no pós mitóticas neurônios), enquanto a indução de mutações de fora do alvo mínimas comparadas com o correspondente mediada por ICLV sistemas de28. Em geral, desenvolvemos um romance, compacto, tudo-em-um kit de ferramentas CRISPR executados em uma plataforma IDLV e descritas as várias vantagens de usar esse veículo entrega para edição avançada do gene.
Aqui, o protocolo de produção do sistema de IDLV-CRISPR/Cas9 é descrito, incluindo as diversas etapas envolvidas em assembly, purificação, concentração e titulação da IDLVs, bem como estratégias para validar a eficácia de gene-edição destes vetores. Este protocolo é facilmente escalável para atender as necessidades de diferentes investigadores e é projetado para gerar com êxito vetores de LV, com uma concentração na faixa de 1 x 1010 transducing unidades (TU) / mL. Os vetores gerados através deste protocolo podem ser utilizados para infectar eficientemente vários diferentes tipos de células, incluindo células-tronco embrionárias difícil-para-transduce, células hematopoiéticas (células T e macrófagos) e culta e vivo em– neurônios injetados. Além disso, o protocolo é igualmente adequado para a produção de vetores de Lentivirus integrase-competente em quantidades semelhantes.
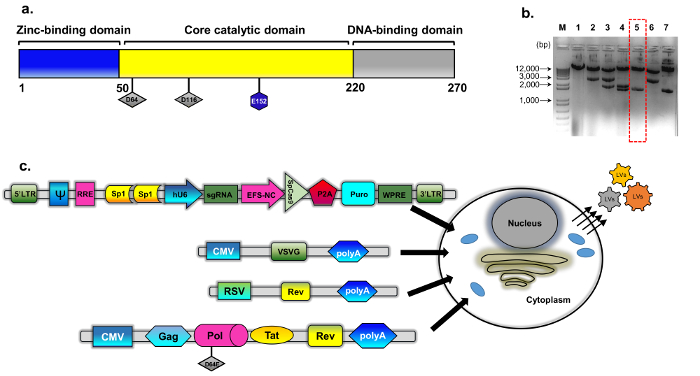
Figura 1: embalagem de IDLV. (a) esquema do tipo selvagem integrase proteína (b) o plasmídeo modificado foi derivado de psPAX2 (ver métodos, construção de plasmídeo para detalhes). Imagem de gel de agarose representante dos clones selecionados para clones mutantes integrase. Preparado usando um isolamento de DNA plasmídeo padrão mini kit de amostras de DNA foram analisadas por digestão com EcoRV e SphI. O clone digerido corretamente (número 5, caixa vermelha tracejada) verificou-se ainda mais por sequenciamento direto (Sanger) para a substituição de D64E em INT. A fita de empacotamento deficiente integrase foi nomeada pBK43. (c) diagrama esquemático do protocolo do transfection transiente empregadas para gerar vetores de IDLV-CRISPR/Cas9, mostrando 293T células transfectadas com VSV-G, embalagens e fitas do transgene (Sp1-CRISPR/Cas9 tudo-em-um plasmídeo). Partículas virais que brote para fora da membrana celular contêm o RNA completo do vetor (expressado o transgene cassete). A segunda geração do sistema de embalagem-IDLV foi usada, que inclui as proteínas reguladoras Tat e expressão de Rev. Rev é complementada ainda mais de uma gaveta separada (RSV-REV-plasmídeo). Abrev: vírus de repetição, VSV-G, estomatite vesicular LTR-Long-terminal G-proteína, promotor de pCMV-citomegalovírus; Promotor de Rous sarcoma vírus (RSV); RRE-(elemento de resposta Rev). Outros elementos reguladores na gaveta a expressão incluem locais Sp1-obrigatórios, elemento Rev Response (RRE), marmota hepatite vírus Posttranscriptional regulamentar elemento (WPRE), um promotor de 1 α fator núcleo-alongamento (EFS-NC), a embalagem de vetor elemento ψ (psi), promotor de citomegalovírus (CMV) humano e humano U6 promotor. Clique aqui para ver uma versão maior desta figura.
