


ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Nörodejenerasyonda nöron-glial etkileşimler yetersiz araç ve yöntemler nedeniyle iyi anlaşılamamıştır. Burada, insan pluripotent kök hücrelerinden indüklenmiş nöronlar, oligodendrosit öncü hücreleri ve oligodendrositler elde etmek için optimize edilmiş protokolleri açıklıyoruz ve Alzheimer hastalığında hücre tipine özgü katkıları anlamada bu yöntemlerin değerlerine örnekler sunuyoruz.
Abstract
Alzheimer hastalığı (AD) ve diğer nörodejeneratif bozukluklarda, oligodendroglial başarısızlık yaygın bir erken patolojik özelliktir, ancak özellikle beynin gri maddesinde hastalık gelişimine ve ilerlemesine nasıl katkıda bulunduğu büyük ölçüde bilinmemektedir. Oligodendrosit soy hücrelerinin işlev bozukluğu, miyelinasyondaki eksiklikler ve oligodendrosit öncü hücrelerinin (OPC'ler) bozulmuş kendini yenilemesi ile işaretlenmiştir. Bu iki kusur, en azından kısmen, patolojinin birikmesi boyunca nöron ve oligodendrositler arasındaki etkileşimlerin bozulmasından kaynaklanır. OPC'ler, CNS gelişimi sırasında miyelinizan oligodendrositlere yol açar. Olgun beyin korteksinde, OPC'ler başlıca proliferatif hücrelerdir (toplam beyin hücrelerinin ~% 5'ini oluşturur) ve yeni miyelin oluşumunu nöral aktiviteye bağımlı bir şekilde kontrol eder. Bu tür nöron-oligodendrosit iletişimi, özellikle AD gibi nörodejeneratif koşullar bağlamında, uygun araçların eksikliği nedeniyle önemli ölçüde az çalışılmaktadır. Son yıllarda, grubumuz ve diğerleri, insan pluripotent kök hücrelerinden bireysel olarak fonksiyonel nöronlar ve oligodendrositler üretmek için mevcut protokolleri geliştirmek için önemli ilerlemeler kaydetmiştir. Bu yazıda, nöron-oligodendrosit bağlantılarını modellemek için bir ko-kültür sisteminin kurulması da dahil olmak üzere optimize edilmiş prosedürlerimizi açıklamaktayız. Açıklayıcı sonuçlarımız, OPC'lerin / oligodendrositlerin beyin amiloidozuna ve sinaps bütünlüğüne beklenmedik bir katkısını önermektedir ve bu metodolojinin AD araştırması için faydasını vurgulamaktadır. Bu indirgemeci yaklaşım, spesifik hetero-hücresel etkileşimleri beynin içindeki doğal karmaşıklıktan ayırmak için güçlü bir araçtır. Burada tanımladığımız protokollerin, nörodejenerasyonun patogenezinde oligodendroglial defektler üzerine gelecekteki çalışmaları kolaylaştırması beklenmektedir.
Introduction
Oligodendrosit soy hücreleri - oligodendrosit öncü hücreleri (OPC'ler), miyelinizan oligodendrositler ve aralarındaki geçiş tipleri dahil - nöral gelişim ve yaşlanma boyunca merkezi sinir sistemimizin düzgün çalışması ve bakımı için birçok kritik fonksiyona aktif olarak katılan önemli bir insan beyin hücresi grubunu oluşturur1 2,3,4 . Oligodendrositler, nöronal aktivite iletimini kolaylaştırmak ve beyaz cevherde aksonal sağlığı desteklemek için miyelin üretmesiyle iyi bilinirken, OPC'ler, miyelinasyonun az olduğu gri maddede bol miktarda (~% 5) bulunur ve öğrenme davranışını ve hafıza oluşumunu yönetmek için aktiviteye bağlı sinyal işlevlerini yerine getirir 5,6,7,8 . Alzheimer hastalığı (AD) ve diğer yaşa bağlı nörodejeneratif durumların patogenezinde oligodendroglial hücrelerin nasıl işlev gördüğü ve işlev bozukluğu az çalışılmıştır9. Uygun bir model sistemin yetersizlikleri ve deneysel bir yolu ileriye götürecek genel bilgi eksiklikleri bu boşluğun başlıca nedenleridir.
İnsan beyin hücrelerinin, embriyonik kök (ES) ve indüklenmiş pluripotent kök (iPS) hücreler de dahil olmak üzere pluripotent kök hücrelerden türetilmesindeki en son atılımların ışığında, modern gen düzenleme araçlarıyla birlikte bu tür hücresel modeller, beyindeki hücresel etkileşimlerin karmaşık bağlantı noktasını ele almak için sağlam araçlar olarak ortaya çıkmıştır ve insana özgü hastalık belirtilerini gösterebilmektedir10, 11. Bireysel beyin hücresi tiplerinin, aynı AD'yi teşvik eden koşullar12,13 karşısında farklı ve hatta çelişkili etkiler gösterebileceği göz önüne alındığında, bu kök hücre metodolojisi, daha önce beyin hücresi tiplerinin koleksiyonlarından yalnızca toplu okumalar sağlayan in vivo veya in vitro modeller kullanılarak gözden kaçırılmış olan hücre tipine özgü bilgileri benzersiz bir şekilde sunar. Son on yılda, ES / iPS hücrelerinin trans-farklılaşmasından veya diğer ölümcül olarak farklılaşmış hücre tiplerinden (örneğin, fibroblastlar) doğrudan dönüşümden insan nöronları üretmek için çok sayıda güvenilir protokol geliştirilmiştir 14,15. Özellikle, anahtar nörojenik transkripsiyon faktörlerinin (örneğin, nörogenin 2, Ngn2)16'nın insan pluripotent kök hücrelerine uygulanması, glial hücrelerle pıhtılaşmaya gerek kalmadan saf kültürler için iyi karakterize edilmiş nöronal hücre tiplerinin homojen bir popülasyonunu oluşturabilir12,17,18. İndüklenmiş insan oligodendrositleri için, birincil meslektaşlarına çok benzeyen fonksiyonel hücreler üretebilen, zaman ve kaynaklarda geniş bir verimlilik ve talep yelpazesine sahip birkaç yayınlanmış protokol vardır 19,20,21,22,23,24,25,26,27,28 . Bugüne kadar, oligodendroglial hücrelerin AD patogenezine nasıl tepki verdiğini ve onu nasıl etkilediğini araştırmak için bu protokollerin hiçbiri uygulanmamıştır.
Burada, insan kaynaklı nöronların (iN'ler) ve OPC'lerin / oligodendrositlerin (iOPC'ler / iOL'ler) tek ve karışık kültürleri için geliştirilmiş protokollerimizi açıklıyoruz. Burada açıklanan iN protokolü, yaygın olarak kullanılan Ngn2 yaklaşımı16'ya dayanmaktadır ve glia-free olma özelliğine sahiptir. Ortaya çıkan iN'ler homojendir ve karakteristik piramidal morfoloji, gen ekspresyon paterni ve elektrofizyolojik özellikler17,18 ile kortikal tabaka 2/3 uyarıcı nöronlara oldukça benzemektedir (Şekil 1). Pluripotent kök hücrelerin yönlendirilmiş farklılaşmasındaki bazı temel engellerin üstesinden gelmek için, düşük doz dimetil sülfoksit (DMSO) ön tedavisi29,30 için basit ve etkili bir yöntem geliştirdik ve Douvaras ve Fossati32 tarafından geniş çapta uyarlanmış bir protokole dayanarak, insan ES / iPS hücrelerinin iOPC'lere ve iOL'lere31'e dönüşme eğiliminin arttığını bildirdik. . Protokolü daha da basitleştirdik ve oligodendroglial olgunlaşma sürecini hızlandırmak için sağlam bir farklılaşmayı teşvik eden bileşik olan clemastine 7,33,34'ü dahil ettik. Sonuç olarak (Şekil 2), iOPC'ler 2 hafta içinde (O4 belirteci için ~% 95 pozitif) ve iOL'ler dört hafta içinde (olgun belirteçler MBP ve PLP1'i ifade eder) üretilebilir. İlginç bir şekilde, iOPC'lerin tek başına kayda değer miktarda amiloid-β (Aβ) salgıladığını, amiloid öncü proteininin (APP) bol miktarda ekspresyonunu ve oligodendrositsoy hücrelerinde proteaz β-sekretazın (BACE1) işlendiğini gösteren bağımsız transkriptomik verilerle tutarlı olduğunu bulduk. Ayrıca, iN-iOPC ko-kültür sistemimiz, aksonların MBP-pozitif iOL süreçleri ile kaplanmasını teşvik eder ve sinaps oluşumu için önemli destek sağlar (Şekil 3). Bu nedenle, aşağıda detaylandırdığımız protokoller, daha önce kataloglanmış nöron-oligodendroglia ortak kültürleme yöntemlerine göre teknik ve biyolojik avantajlara sahiptir ve AD'de nörodejenerasyonun daha iyi modellenmesinde bir umut vaat etmektedir.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. İnsan pluripotent kök hücrelerinden insan nöron indüksiyonu
- Lentivirüs hazırlığı (~ 5 gün, daha önce açıklandığı gibi ayrıntılı protokol16)
- Her T75 şişesinde ~1 milyon HEK293T hücresi plakası, transfeksiyon yaparken ~% 40 oranında bir araya gelmelerini sağlar. Tetrasiklin ile indüklenebilir Ngn2 ve püromisine dirençli geni (PuroR; aynı TetO promotör kontrolü altında), rtTA ve üç yardımcı plazmid pRSV-REV, pMDLg / pRRE ve VSV-G'yi (12 μg lentiviral vektör DNA'sı ve yardımcı plazmid DNA'sının her birinin 6 μg'ı) eksprese eden plazmidlerle transfekte edin. Lentivirüs preparatı başına en az üç şişe hazırlayın. Üreticinin talimatını izleyerek transfeksiyon için PEI kullanın. Medyayı 16 saat sonra değiştirin ve atın.
- Hasat, her gün kültür medyasını toplayarak viral parçacıkları serbest bıraktı ve 3 gün boyunca taze medya ile değiştirin. Toplanan medyayı saflaştırma için viral parçacıklar içeren havuzlayın. Virüsü 0,22 μm'lik bir filtreden geçirin ve 90 dakika boyunca 49.000 x g'de santrifüj yapın. Peleti uygun PBS-glikoz hacminde (~ 150 μL) yeniden askıya alın.
- Nöron İndüksiyonu (~ 5 gün)
NOT: Bu indüksiyon protokolü (Şekil 1A; akış diyagramı), doğrulanmış pluripotensi olan hem iPS hem de ES hücreleri için oldukça etkilidir (iyi karakterize edilmiş pluripotens belirteçlerinin immünohistokimya boyaması ile test edilebilir; Şekil 1B).- Ticari olarak temin edilebilen H1 insan ES hücrelerini 52'nin geçişinde kullanın (bakınız Malzeme Tablosu). ES hücre bakım ortamı (bakınız Malzeme Tablosu) kullanarak hücre dışı matris çözeltisi kaplamalı 6 delikli plakalar (6 delikli plaka başına ~ 0.5 mg matris çözeltisi; bakınız Malzeme Tablosu) üzerindeki hücreleri kültürleyin ve plakaları% 5 CO2 ile 37 ° C'de inkübe edin.
- -2. Günde, ES hücrelerini (% 80 birleşik) 1 mL hücre ayırma çözeltisi ile ayırın ( Malzeme Tablosuna bakınız) ve 10 dakika boyunca oda sıcaklığında inkübe edin. Hücreleri bir tüpe aktarın; kuyuyu 2 mL medya ile yıkayın ve aynı tüpte birleştirin. 5 dakika boyunca 300 x g'de santrifüj yapın, peleti ortama geri koyun ve hücreleri, kuyu başına 1 x 105 hücrelik tohumlama yoğunluğunda matris kaplı 6 kuyucuklu plakalara yerleştirin.
- -1. Günde, taze ES hücre bakım ortamındaki ES hücrelerine polibren (8 μg / ml) ile birlikte Ngn2 artı PuroR ve rtTA'yı eksprese eden lentivirüsleri ekleyin (bkz. Tam virüs miktarı, gerçek titreler veya titrasyon ile belirlenmelidir. Genellikle 6 delikli bir plakaya kuyucuk başına her virüs 5 μL ekleriz.
- 0. Günde, DMEM-F12 ortamında morfojensiz N2 takviyesi ile Doksisiklin (2 μg / mL, Ngn2 ekspresyonunu aktive etmek için) ekleyin.
- 1. Günde, taze DMEM-F12 artı N2 ve doksisiklin ortamında, 1 μg / mL ortamın son konsantrasyonuna Puromycin ekleyin. Virüs titresi düşükse, az transdücin hücreleri yeterince çıkarmak için daha yüksek Puromisin konsantrasyonu (5 μg / mL'ye kadar) ve daha uzun seçim süresi (48 saate kadar) gerekebilir.
- 2. Günde, farklılaştırıcı nöronları hücre ayrılma çözeltisi ile ayırın (Malzeme Tablosuna bakınız) ve matris çözeltisi ile kaplanmış 24 delikli plakalara (80.000-200.000 hücre / kuyu arasında) yeniden plakalayın ( Malzeme Tablosuna bakınız) ve bunları doksisiklin olmadan NBA / B27 ortamında tutun. Tohumlama yoğunluğu kritiktir.
- Bu aşamada, ayrılmış nöronlar özel ticari dondurma ortamında dondurulabilir ( Malzeme Tablosuna bakınız) ve 3 aya kadar sıvı azotta saklanabilir. Saf nöronlar, çözülme sonrası tipik ~% 15-% 20 hücre ölümünü hesaba katarak, tek başına kültürlenmiş veya diğer beyin hücresi tipleriyle birlikte kültürlenmiş olarak kaplanabilir (OPC'lerle birlikte kültürleme için bkz. adım 3.2.3).
- Üretici tarafından talimat verildiği gibi hücre dışı matris tabanlı çözeltilerle kaplanmış plakalar üzerinde saf iN'leri kültürleyin (bkz. Karakteristik piramidal morfoloji, 4. Gün (ve 6. Gün; Şekil 1C). Sinaps oluşumu 14. ila 16. günler arasında erken tespit edilebilir ve 24. günde standart sinaptik öncesi ve sonrası belirteçlerle immünohistokimyasal boyama ile belirgindir. (Şekil 1D; sinaptik öncesi belirteç Synapsin 1 ve dendritik işaretleyici Map2 ile etiketlenmiştir).
2. Pluripotent kök hücrelerden insan oligodendrosit öncü hücre (OPC'ler) indüksiyonu ve oligodendrosit olgunlaşması
- Nöral Progenitör Hücre (NPC) üretimi: tek katmanlı protokol (~ 7 gün). Akış diyagramı için Şekil 2A'ya bakın.
- Kültür H1 insan ES hücreleri daha önce tarif edildiği gibi (bkz. adım 1.2.1.) ve çoklu sinyal yolları için küçük molekül inhibitörleri ile ikili SMADi adı verilen yerleşik bir yaklaşımla onları nöral progenitör hücrelere (NPC'ler) trans-farklılaştırır. Burada yaygın olarak kabul gören bir ticari kit kullanıyoruz ve üretici tarafından sağlanan tek katmanlı protokolü takip ediyoruz (bkz.
- -1. Günde, ES hücre bakım ortamı ile bir büyüme faktörü azaltılmış matris çözeltisi ile kaplanmış 6 delikli bir plakada kuyucuk başına 0,5–1 x 106 hücre plakası ( bkz. Bu büyüme faktörü azaltılmış matris çözeltisi, aşağıdaki adımlarda kullanılacak tüm plakaları kaplamak için kullanılır.
- 0. Günde, hücreleri% 2 DMSO ile desteklenen ES hücre bakım ortamı ( bakınız Malzeme Tablosu) ile 24 saat boyunca tedavi edin.
- 1-6. Günde, ticari kitten SMAD inhibitörlerini içeren sıcak (37 ° C) nöral indüksiyon ortamı ile tam ortamı değiştirin (bkz. Hücreler 7. Gün'den önce bölünür ve birleşime ulaşırsa, onları adım 2.1.2'de daha önce açıklandığı gibi 0.5-1 x 106 tohumlama yoğunluğuna geçirin.
- 7. Günde, hücre ayrılma çözeltisi (bakınız Malzeme Tablosu) ve 24 delikli bir plakanın 1-2 x 105 hücreli / kuyucuklu bir tohumlama yoğunluğunda plaka kullanarak NPC'leri geçiş.
- İmmünohistokimyasal (IHC) boyama ile pluripotens belirtecinin yokluğu, örneğin OCT4 ve PAX6, Nestin ve Sox1 gibi NPC belirteçlerinin varlığı için farklılaşma etkinliğini test edin.
- Bu aşamada, ayrılmış NPC'ler özel ticari NPC dondurma ortamında dondurulabilir ( Malzeme Tablosuna bakınız) ve 3 aya kadar sıvı azotta saklanabilir. Bir kez dondurulup çözüldükten sonra, NPC'ler hala güvenilir protokollere sahip nöronlara, astrositlere ve OPC'lere yol açmak için çok potansiyeli korurlar.
- Oligodendrosit öncü hücre (OPC) üretimi (~ 7 gün). Akış şeması için lütfen Şekil 2A'ya bakınız.
- 7. Günde, hücre ayrılma çözeltisi kullanarak NPC'leri pasaj yapın (bakınız Malzeme Tablosu) ve bunları sıcak (37 ° C) nöral indüksiyon ortamında ve ticari kitten SMAD inhibitörlerinde 24 delikli bir plakada kuyucuk başına 1-2 x 105 hücrelik bir tohumlama yoğunluğunda plakalayın (bkz.
- 8. Günde, OPC farklılaşma ortamında% 1'lik bir DMSO çözeltisi hazırlayın ve kaplanmış NPC'leri 24 saat boyunca tedavi edin. OPC farklılaşma ortamı şunlardan oluşur: DMEM/F12 ortamı, %1 N2 takviyesi, %1 B27 takviyesi, 20 ng/mL'de bFGF, 1 μM'DE SOG, 10 ng/mL'de PDG-AA (bkz.
- 9. Günde, medyayı DMSO içermeyen yeni OPC farklılaştırma ortamıyla değiştirin. Hücreleri 15. Güne kadar her gün besleyin. Hücreler 15. Gün'den önce birleşime ulaşırsa, bunları adım 2.2.1'de açıklandığı gibi 1-2 x 105 hücrenin tohumlama yoğunluğuna geçirin.
- 14. Günde, OPC farklılaşma ortamındaki plaka OPC'leri, 24 delikli bir plakada 1–2 x 105 hücre/kuyu yoğunluğunda ortalanır.
- Bu aşamada (15. Gün), hücreleri IHC boyama veya qPCR ile OPC'ye özgü belirteçlerin varlığı açısından test edin (örneğin, O4, Olig1/2, CSPG4/Ng2, NKX2.2, PDGFRa; Şekil 2B) ve NPC belirteçlerinin yokluğu için (Pax6 veya Nestin; Şekil 2D). Tipik olarak O4 immünoreaktivitesini 15. günde hücrelerin% 95'inden fazlasında tespit ediyoruz. Alzheimer hastalığı ile özellikle ilgili olarak, APP (amiloid öncü proteini), BACE1 (işleme proteaz β-sekreataz 1) ve peptid amiloid-β (Aβ) ekspresyonu OPC'lerde bol miktarda bulunur (Şekil 2F).
- Oligodendrosit (OL) olgunlaşması (~7–20 gün)
- 15. Günde, ortamı OL olgunlaşma ortamı ile değiştirin: Nörobazal-A ortamı,% 2 B27 takviyesi, 1 μM cAMP, 200 ng / mL T3 triiodotironin ve 1 μM'lik Clemastine (bkz. Gerekirse ortamı her gün veya her gün değiştirin.
- Hücreler% 90 birleşime ulaştığında, 2 pasaja kadar 1: 3 oranında veya hücre bölünmesi önemli ölçüde yavaşlayana kadar bölünür. OPC'ler çok hızlı bölünürse ve 3 günden daha kısa sürede akıcılığa ulaşırsa, 1-3 gün boyunca 2-5 μM'lik bir konsantrasyonda Ara-C'yi ( Malzeme Tablosuna bakınız) ekleyin. Aktif proliferasyon, olgunlaşma verimliliğinin azaldığını gösterir.
- OL belirteçlerinin ekspresyonunu değerlendirerek oligodendroglial olgunlaşmanın etkinliğini inceleyin, örneğin, CLDN11, PLP1, MBP, qPCR, IHC boyama veya immünoblotlama ile. Son derece karmaşık yapıların karakteristik morfolojisi (Şekil 2C) ve OL belirteçlerinin ekspresyonu (Şekil 2E) 28. Güne kadar kolayca tespit edilmelidir.
3. İnsan kaynaklı nöronların (iN'ler) ve oligodendrosit öncü hücrelerinin (iOPC'ler) birlikte kültürlenmesi
- iOPC kaplama (~3 gün)
- OPC farklılaştırma ortamında (adım 2.2.2'de açıklandığı gibi) 24 delikli bir plakada (yukarıda adım 2.2.4'te açıklandığı gibi) kuyu başına 1 x 105 hücre yoğunluğunda 14. Gündeki plaka iOPC'leri.
- iN-iOPC ortak kültür kurulumu
- 15. Günde, indüklenen insan nöronlarını, Puromisin seçiminden sonraki 2. Gün adımında (adım 1.2.6'da açıklandığı gibi) hücre ayrılma çözeltisi ile ayırın (bkz.
- Kültürlenmiş OPC'lere nöronlar ekleyin, büyüyen OPC'lerle (adım 3.1.1'den) 24 delikli plakada kuyu başına 2 x 105 hücrenin tohumlama yoğunluğunda kaplanın. Nörobazal-A ortamı,% 2 B27 takviyesi ve 100 ng / mL T3 triiodotironin içeren ko-kültür ortamını kullanın. Ortamı ertesi gün ve daha sonra her gün değiştirin. OPC'ler çok hızlı çoğalırsa ve 3 günden daha kısa sürede akıcılığa ulaşırsa, 2-5 μM'lik bir konsantrasyonda Ara-C ekleyin. 7 gün sonra ko-kültürde yetiştirilen iNs ve iOPC'lerin temsili bir görüntüsü Şekil 3A'da gösterilmiştir.
- OPC'lerle birlikte kültürleme için yukarıda adım 1.2.7'de açıklandığı gibi hazırlanmış dondurulmuş nöronları kullanın. Plaka donma ve çözülme nöronları, kuyu başına 3 x 105 hücrenin daha yüksek bir yoğunluğunda.
- Ko-kültürlerde 14-16. Gün'den sonra, iN'lerdeki sinaps oluşumu, sinaptik belirteçlerin IHC boyaması ile gözlemlenebilir ve 21. Gün'e kadar sinaptik punkta bol olmalıdır (Şekil 3C) ve nöronal aktiviteler güvenilir bir şekilde kaydedilebilir.
- 21. Gün'den itibaren, hücreleri OL'ye özgü belirteçler (örneğin, MBP ve PLP1) için test edin. 28. Günde, normalde iN aksonlarının iOL süreçleri tarafından kaplanması fenomenini gözlemleriz, spesifik belirteçler için IHC boyaması ile etiketlenir (Şekil 3B; iN aksonları için nörofilament NF ve iOPC süreçleri için MBP).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
İnsan pluripotent kök hücrelerinden insan kaynaklı nöronların doğrudan üretimi
Yeni başlayan insan pluripotent kök hücrelerinin, başarılı bir şekilde iN veya iOPC / iOL üretimi için yüksek derecede pluripotens sergilemesi çok önemlidir. Bu nedenle, hücreler bu makalede açıklanan indüksiyon protokollerinden herhangi birine başlamadan önce Oct4 ve SOX2 gibi spesifik belirteçler için boyanmalıdır (Şekil 1A). İnsan H1 hücreleri, Zhang ve ark. tarafından daha önce yayınlanan protokolü takiben indüklenmiş uyarıcı ön beyin nöronlarını elde etmek için kullanıldı. bazı modifikasyonlarla (Şekil 1C) 12,16,17,18. Burada, 2. Gündeki iN'lerin, herhangi bir besleyici tabakanın yokluğunda, matris çözeltisi üzerinde saf kültürde yeniden kaplandığı bir protokol sunuyoruz (bakınız Malzeme Tablosu). glia veya fibroblastlar. Daha önce yayınlanan protokollere ek olarak, 2. günde iN'lerin dondurulmasının hücre canlılığını önemli ölçüde etkilemediğini gözlemliyoruz (çözülmeden sonra ~% 15-20 hücre ölümü). Kültürdeki saf nöronlar 14-16. günlerde sinapsin1'i ifade etmeye başlayacaktır (Şekil 1D). Saf bir nöronal kültür oluşturmak çok önemlidir, çünkü bazı faktörler, örneğin, önde gelen AD risk faktörü ApoE, besleyici tabakadaki hücreler tarafından eksprese edilebilir ve bu da sonuçları önemli ölçüde karıştırabilir.
iOPC üretimi ve iOL olgunlaşması DMSO tedavisi ile iyileştirilir
Burada, iOPC'lerin 2 haftada ve olgun iOL'lerin 4-5 haftada üretilmesini sağlayan hızlı ve verimli bir protokol sunuyoruz (Şekil 2A). ES ve iPS hücreleri29,30,31 için farklılaşma verimliliğini artırmak için daha önce geliştirdiğimiz geçici DMSO tedavisi yönteminden yararlandık. DMSO tedavisi, daha iyi sinyal entegrasyonu için erken G1 fazındaki hücre sayısını zenginleştirir ve farklılaşmayı destekler. İnsan ES hücrelerini NPC'ler üretmeye teşvik etmeden önce ilk tedaviyi ve NPC'leri iOPC'lere ayırmadan önce ikinci tedaviyi gerçekleştirdik. Spesifik OPC belirteçlerini (Olig2, CSPG4, NKX2.2 ve PDGFRA) ES H1 hücrelerinin kaplanmasından 2 hafta sonra tespit edebiliriz (Şekil 2B, E). Bu aşamadaki iOPC popülasyonu oldukça homojendir, hücrelerin %>95'i O4 boyaması için pozitif ve diğer belirteçler için yüksek düzeyde immünoreaktivite (Şekil 2B). 15. Günde OL olgunlaşmasının başlamasından sonra, tipik olarak 28. Gün'den itibaren belirli OL belirteçlerini (MBP, O1, CLDN11 ve PLP1) tespit edebiliriz (Şekil 2C, E). Bu aşamaya özgü belirteçlerin ekspresyonu, oligodendroglial hücrelerin gelişimsel seyri ile ilişkilidir ve NPC belirteçlerinin giderek azaldığı, OPC belirteçlerinin ikinci hafta civarında zirveye ulaştığı ve OL belirteçlerinin üçüncü haftada yükseldiği hızlandırılmış bir hıza işaret eder (Şekil 2D, E)37. Bu olgunlaşma sürecinin hücre popülasyonlarını çeşitlendirdiğini lütfen unutmayın. OPC'ler ve olgun miyelinizan oligodendrositler arasındaki çoklu ara aşamalardan oluşan süreklilikteki alt popülasyonlar mevcut olabilir ve daha sonra daha olgun hücrelerin baskın olduğu toplam hücrelerin değişen yüzdesini oluşturabilir.
Bir karşılaştırma olarak, yüksek referanslı iOPC'leri satın aldık ve üretici protokolünü izleyerek iOL'de olgunlaştırdık. Yukarıda belirtilen belirteçlerin ekspresyonunu hem iOPC hem de iOL preparatlarımızda ve satın aldığımız hücrelerde test ettik. Protokolümüzü takiben üretilen hücrelerin, test edilen tüm genlerin daha yüksek ekspresyonuna sahip olduğunu belirledik (Şekil 2E). İlginç bir şekilde, iN'lerde iOPC'lere karşı iki ana amiloid-β izoformunun (Aβ40 ve Aβ42) salgılanan seviyelerini test ettiğimizde, iOPC'lerin her iki parçadan da daha fazlasını salgıladığını, ancak oranın aynı kaldığını fark ettik (Şekil 2F).
iNs ve iOPC'lerin birlikte kültürlenmesi
Bu protokol, özellikle iN'leri ve iOPC'leri birlikte kültürlemek için optimize edilmiştir ve sinirsel gelişim boyunca bu iki hücre tipi arasındaki hücreler arası iletişimin gerçek zamanlı olarak izlenmesini sağlar. Her iki hücre tipi için ideal kaplama yoğunluklarına, uygun farklılaşmayı sağlamak için bir dizi hücre numarası titrasyonu ile karar verilmelidir (Şekil 3A). Ko-kültürlerde 4 hafta sonra, iOPC'lerin MBP gibi spesifik belirteçler için pozitif olan OL'lara yeterince farklılaşması ve süreçleri ensheath aksonlarına genişletmesi beklenmektedir (Şekil 3B). Ko-kültür sistemi, sinaps sayısını sağlam bir şekilde artırabilir, bu da iOPC'lerin fiziksel temaslar veya trofik faktörlerin salınması yoluyla nöronal bir destek sağladığını gösterir (Şekil 3C). Ko-kültürleri 6 haftaya kadar kabul edilebilir sağlık durumunda tutabilir ve sinaps sayısının ve diğer nöronal özelliklerin beşinci hafta civarında plato oluşturduğunu gözlemleyebiliriz. Astrositler ve mikroglia preparatlarımızda mevcut değildir ve bunların yokluğu, belirli belirteçlerin ekspresyonu kontrol edilerek belgelenebilir (Şekil 3D). iOPC'ler, komşu nöronlardan gelen aktiviteye bağımlı sinyallere potansiyel olarak yanıt verebilecek ve aracılık edebilen, parakrin (örneğin nörotrofinler ve metabolitler) ve / veya sinaptik bir şekilde (Şekil 3E ve 3F) çok sayıda iyi karakterize edilmiş geni ifade eder.

Şekil 1: hPSC'lerden insan kaynaklı nöronların (iN) doğrudan üretimi. (A) iN üretiminin akış diyagramı. (B) Pluripotensi doğrulamak için insan pluripotent kök hücrelerinin (H1) başlangıç kültürünün temsili parlak alan ve immünofloresan görüntüleri. Oct4 kırmızı ve Sox2 yeşil renkte gösterilir. (C) 4. Gün ve 6. Gündeki iN'lerin temsili parlak alan görüntüleri. (D) 24 gün boyunca saf kültürde yetiştirilen ve dendritik belirteç Map2 ve pre-sinaptik belirteç Synapsin 1 (Syn1) için immünofloresan boyama ile boyanan iN'lerde dendritik arborizasyon ve sinaps punkta için karakteristik morfoloji. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.

Şekil 2: iOPC oluşturma ve iOL olgunlaşması. (A) iOPC ve iOL üretiminin akış diyagramı. (B) 15. Günde iOPC'lerin temsili parlak alan ve immünofloresan görüntüleri. Olig2 (pan-oligodendroglia marker) yeşil, O4 (OPC marker) kırmızı ve DAPI mavi renkte gösterilir. Görüntülemede iOPC'lerin %>95'inin O4 ve %25'inin Olig2 için pozitif olduğu saptandı. (C) 28. Günde iOL'lerin temsili parlak alan ve immünofloresan görüntüleri. MBP yeşil, O1 kırmızı ve DAPI mavi renkte gösterilir. (D) NPC belirteci PAX6'nın ekspresyonu, 14. Günde iOPC'lerde çarpıcı bir şekilde azalır ve 28. Gün'de OL'lerde arka plana daha da düşer, bu da güçlü bir NPC trans-farklılaşmasına ve iOPC popülasyonunda yüksek düzeyde homojenliğe işaret eder. (E) Tanımlanan protokol tarafından üretilen kültürlerdeki ortak OPC ve OL belirteç genlerinin zaman seyri ekspresyon profili, DMSO inkübasyonunun adımı (adım 2.1.3 ve 2.2.2) olmadan (-DMSO) veya (+DMSO) farklı zaman noktalarında test edilmiştir. Karşılaştırma olarak, ticari iOPC'ler ( Malzeme Tablosuna bakınız) üreticinin talimatlarına göre olgunlaştırıldı ve hem iOPC'ler (iOPC-Tempo) hem de iOL'ler (iOL-Tempo) aynı belirteçler için test edildi. Beklendiği gibi, MBP (olgun bir oligodendrosit belirteci), test edilen tüm iOPC'lerde farklılaşmanın erken aşamalarında tespit edilmemiştir (N.D.). DMSO, OPC farklılaşmasının ve OL olgunlaşmasının verimliliğini önemli ölçüde artırdı. (F) Aβ40 ve Aβ42'nin saf iNs ve iOPC kültürlerinde üretilmesi ve salgılanması, hem 15. günde hem de hücre numaralarıyla normalleştirilen (her ikisi de 24 delikli bir plakada kuyu başına 200.000 hücre yoğunluğunda) ticari ELISA kitleri ( Malzeme Tablosuna bakınız) ile ölçülür. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Çubuk grafiklerdeki veriler ortalama SEM ± (n ≥ 3) olarak çizilir. İstatistiksel anlamlılık öğrencinin t-testi ile değerlendirildi (*, p < 0.05; ***, p < 0.001); NPC ile karşılaştırıldığında (D) içinde; in (E), iOPC-Tempo kontrolüne kıyasla; in (F), iN ile karşılaştırıldığında.

Şekil 3: iN'lerin ve iOPC'lerin ortak kültürü. (A) 7. Gündeki ko-kültürlü iN'lerin ve iOPC'lerin temsili parlak alan görüntüsü, daha fazla olgunlaşma için uygun bir yoğunluk gösterir. (B) 28 gün boyunca birlikte kültürlenmiş iNs ve iOPC'lerin temsili immünofloresan görüntüsü. Aksonal belirteç nörofilament NF yeşil renkte, oligodendrositik belirteç MBP ise kırmızı renkte gösterilmiştir. Sağda, iOL işlemi (MBP +) tarafından kaplanmış bir iN akson segmenti. (C) 4 haftalık ortak kültürlerde analiz edilen sinaps oluşumu. Hücreler Synapsin 1 (Syn1, yeşil) ve MAP2 (kırmızı) için boyandı ve sinaptik punkta, 17,18'de tanımlandığı gibi dendritik segmentler boyunca yoğunluğun konfokal analizi ile ölçüldü. (D) iNs ve iOPC'lerin ortak kültürlerinde (7 günlük birlikte kültürleme), astrosit belirteçlerinin, ALDHL1 ve GFAP'nin ekspresyonu minimumdur (üstte) ve mikroglia belirteçlerinin, TMEM119, TREM2 ve CD33'ün ekspresyonu, qPCR tarafından tespit edilmez (N.D.). Böylece bu iki glial hücre tipinden kaynaklanan kontaminasyon dışlanır. (E) iOPC'nin iN ile birlikte kültürlenmesi, nöron-OPC sinapslarının oluşumuna yol açar. Floresan etiketli post-sinaptik belirteç PSD95-mCherry sadece OPC'lerde ifade edilir ve tek kültürlerde (solda) dağınık bir desen gösterir, ancak kokültürlerde puncta oluşturmak için agrega gösterir (sağda, oklarla gösterilir; Tuj1, nöronal belirteç). (F) 14. Gün'de iOPC'lerin saf kültürlerinde nöronal aktiviteleri algılayabilen ve bunlara cevap verebilen iyi karakterize edilmiş oligodendroglial genlerin ekspresyonu. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Çubuk grafiklerdeki veriler ortalama ± SEM (n ≥ 3) olarak çizilir. İstatistiksel anlamlılık öğrencinin t-testi ile değerlendirildi (**, p < 0.005; ***, p < 0.001); (C) içinde, OPC koşulu yok; (D), üst paneldeki birincil astrositlerle karşılaştırıldığında.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Sinaps yapılarını stabilize etmek ve miyelinasyon yoluyla tuzlayıcı sinyal iletimini kolaylaştırmak için fiziksel ve metabolik desteğe ek olarak, oligodendrosit soy hücreleri, nöronlarla hızlı ve dinamik çapraz görüşmeler yoluyla nöronal aktivite paternini şekillendirebilir 5,6,7. AD patolojisinde oligodendroglial yanıtlar başlangıçta inflamasyon ve oksidatif streslere ikincil olarak kabul edilirken, şimdi tehlikeye giren miyelin bütünlüğünün Aβ agregasyonu ve tau hiperfosforilasyonunun ortaya çıkmasından önce erken patojenik bir olay olduğunu savunan umut verici kanıtlar vardır9. Ayrıca, OPC'lerin kendi kendini yenilemesi yoluyla miyelinasyonun onarımı, büyük ölçüde nöronal aktivitelere bağlı bir süreç olan AD38'de özellikle savunmasızdır. Sağlıklı nöron-oligodendrosit sinyallemesini destekleme mekanizmasını anlamak, böylece yeni terapötik hedeflerin belirlenmesi için mükemmel bir fırsat sunmaktadır.
Tek transkripsiyon faktörü Ngn2 protokolü, kök hücre kaynaklı insan nöronlarının üretimi için en çok kullanılan tekniklerden biridir ve burada özetlenen prosedürler, saf nöronal kültürlerin elde edilmesi için daha fazla iyileştirmedir. iOPC / iOL protokolümüz, daha önce yayınlanmış çalışmalardan (4 ila 24 hafta) daha kısa bir indüksiyon süresine sahiptir ve yaygın olarak kullanılan diğer protokollerle karşılaştırılabilir sağlam bir verim ve saflığa sahiptir 19,20,22,23,24,25,26,27,28 . Protokolümüz, ES hücrelerinin NPC'lere, OPC'lere ve son olarak oligodendrositlere kademeli olarak farklılaşmasını karakterize edici modelleme ipuçlarıyla tanıtır ve miyelinasyon homeostazının düzenlenmesini incelemek ve önceki çalışmada açıklandığı gibi in vitro veya in vivo (örneğin, titreyen fare modeline aşılama yoluyla) onarımı için kullanılabilecek fonksiyonel hücreler üretir. Protokolümüzdeki iyileşme, retinoblastom proteinini aktive eden ve yönlendirilmiş farklılaşmanın uyaranlarını daha iyi entegre etmek için hücre döngüsünün G1 fazını uzatan ve ayrıca fonksiyonel türevlere terminal farklılaşmasını artıran DMSO inkübasyonu tarafından büyük ölçüde desteklenmektedir29,30. Son olarak, remiyelinasyon terapötikleri33 için ilaç taramaları yoluyla tanımlanan muskarinik ve antihistaminik bir bileşik olan klemastinin kullanımı, iPS hücre hazırlığı ve canlı hayvanlar 7,28'de gözlemlendiği gibi oligodendrosit olgunlaşmasını kısaltır.
Tekniğin sınırlamaları esas olarak basitleştirilmiş in vitro ortamlar ile beyindeki in vivo ortamlar arasındaki içsel tutarsızlıkta yatmaktadır; Bu tutarsızlık, bireysel beyin hücresi tipleri için ileri aşamalarda tam gelişimsel potansiyelde bir azalmaya yol açar. iN'ler için, son çalışmalar kültürleri oldukça uzun bir süre boyunca iyi sinaptik sağlıkta tutabilmiştir, ancak yine de "eski" iN kültürlerinde (25 aylık kültürlerde bile) azalmış omurga benzeri yapılar ve bozulmuş spontan sinaptik iletim olarak ortaya çıkan bazı göreceli olgunlaşmamışlıkları ortaya koymuştur39. İOPC'lerin transgenik fare beyinlerine nakledildikten sonra aksonları in vivo olarak miyelinleştirdiği tekrarlanabilir bir şekilde gösterilmiş olsa da, elektron mikroskobik değerlendirmeli in vitro miyelinasyon testleri, bu protokol de dahil olmak üzere neredeyse tüm yayınlanmış protokoller19,28 için tatmin edici olmayan verimlilikle teknik bir zorluğu temsil etmektedir. Bu nedenle, nöron-OPC ortak kültür sistemimizin, AD patolojisinin geç evresinin yanı sıra beyin yaşlanma sürecini de sadık bir şekilde taklit etmesi beklenmemektedir. Daha ziyade, nöronlar ve OPC'ler veya erken evre oligodendrosit arasındaki ayrıntılı hücreler arası etkileşimleri, miyelinasyondan bağımsız olan ve yine de uygun nöral gelişim ve hastalık patogenezi için temel olan ayrıntılı hücreler arası etkileşimleri çözmek için benzersiz bir şekilde hazırlanmıştır.
Burada açıklanan üç yordamın her birinin protokol içinde kendi c ritical adımları vardır ve değişiklikler ve sorun giderme gerektirebilir. İN protokolü için iki kritik adım vardır: puromisin seçimi (adım 1.2.5) ve kaplama yoğunluğu (adım 1.2.6). Az transdüke olmuş hücrenin eksik çıkarılması, zayıf farklılaşmış hücrelerin kontaminasyonuna neden olur ve nöronal sağkalımı ve fonksiyonları tehlikeye atar. Adım 1.2.5'te açıklandığı gibi daha yüksek konsantrasyon ve daha uzun inkübasyon ile daha güçlü puromisin seçimi için modifikasyonlar dikkate alınmalıdır. Düşük yoğunluk kültürlerin çökmesine yol açtığından ve yüksek yoğunluk hücre agregasyonunu teşvik ettiğinden ve nöronal büyümeyi engellediğinden, uygun kaplama yoğunluğu her pluripotent hücre hattı için titrasyon ile belirlenmelidir. iOPC / iOL protokolü için iki kritik adım, OPC farklılaşmasında hücre çoğalmasının kontrolü (adım 2.2.3) ve OL olgunlaşması için kaplama yoğunluğudur (adım 2.2.4 ve 2.3.2). Farklılaşan NPC'lerin aşırı büyümesi, OPC farklılaşma uyaranlarına zayıf bir yanıt sinyali verir ve uygun bir Ara-C tedavisi dozu (belirtilen aralık içinde) ile sönümlenmesi gerekir. OPC'leri olgunlaşma için kaplarken, seyrek dağılım karmaşık yapıların fizyolojik morfolojisinin indüksiyonunu kolaylaştırabileceğinden burada daha düşük bir hücre yoğunluğu aralığı tercih edilir ( Şekil 2C'de gösterildiği gibi). iN-iOPC ortak kültürleme protokolü için, her iki hücre tipi için de uygun yoğunlukta kaplamanın kritik adımına dikkat çekmek istiyoruz (adım 3.1.1 ve 3.2.2). Spesifik olarak, iN'ler büyüyen OPC'ler arasında yüzeye iyi yapışmayabilir ve kültür akıcılığa ulaştığında ilk önce ayrılma eğilimindedir. En uygun orana, hücre sayılarının titre edilmesiyle karar verilmelidir.
Genel olarak, protokollerimizde bulunan bu indirgemeci yaklaşım, spesifik hetero-hücresel etkileşimleri insan beyninin doğal karmaşıklığından ayırmak ve sağlıkta ve AD'de oligodendroglial biyolojiyi ortaya çıkarmaya hizmet etmek için güçlü bir araçtır. Mevcut yöntemlere göre önemi bu nedenle bize göre oldukça açıktır. Gelecekteki uygulamalar için burada geliştirilen yöntemlerin ek bir faydası, radyoterapi sonrası 40 ve omurilik yaralanması41,42 gibi demiyelinizan durumlar için hücre bazlı tedavidir. Dahası, bu kök hücre tabanlı sistemin yüksek verim kapasitesi, nöronların, OPC'lerin, oligodendrositlerin fizyolojik durumunu ve etkileşimlerini koruyabilen veya geri yükleyebilen bileşikler için küçük moleküllerin kütüphanelerini taramak için daha büyük ölçekte de kullanılabilir. Bu nedenle, burada açıklanan protokollerin, AD ve diğer nörodejeneratif bozukluklar için daha iyi modelleme araçları ve etkili tedaviler geliştirmede gelecekteki çalışmaları kolaylaştıracağına inanıyoruz.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Yazarların açıklayacak hiçbir şeyleri yoktur.
Acknowledgments
Bu çalışma, Ulusal Sağlık Enstitüleri (R00 AG054616'dan Y.A.H.'ye ve T32 GM136566'dan K.C.'ye), Stanford Üniversitesi Tıp Fakültesi ve Siebel Bursu'ndan (S.C.'ye verildi) gelen hibelerle desteklenmiştir. Y.A.H., Brown Translasyonel Bilimler Enstitüsü'ndeki Translasyonel Sinirbilim Merkezi'nden GFL Translasyonel Profesörüdür.
Materials
| Name | Company | Catalog Number | Comments |
| Accutase | STEMCELL Technologies | 7920 | |
| B27 supplement | ThermoFisher | 17504044 | |
| bFGF | ThermoFisher | PHG 0266 | |
| cAMP | MilliporeSigma | A9501 | |
| Clemastine | MilliporeSigma | SML0445 | |
| DMEM/F12 medium | STEMCELL Technologies | 36254 | |
| DMSO | ThermoFisher | D12345 | |
| Doxycycline | MilliporeSigma | D3072 | |
| Fetal Bovine Serum | ScienCell | 10 | |
| H1 human ES cells | WiCell | WA01 | |
| Matrigel | Corning | 354234 | |
| mTeSR plus | STEMCELL Technologies | 5825 | |
| N2 supplement | ThermoFisher | 17502001 | |
| Neurobasal A medium | ThermoFisher | 10888-022 | |
| Non Essential Amino Acids | ThermoFisher | 11140-050 | |
| PDGF-AA | R&D Systems | 221-AA-010 | |
| PEI | VWR | 71002-812 | |
| pMDLg/pRRE | Addgene | 12251 | |
| Polybrene | MilliporeSigma | TR-1003-G | |
| pRSV-REV | Addgene | 12253 | |
| Puromycin | ThermoFisher | A1113803 | |
| ROCK Inhibitor Y-27632 | STEMCELL Technologies | 72302 | |
| SAG | Tocris | 4366 | |
| STEMdiff Neural Progenitor Freezing Media | STEMCELL Technologies | 5838 | |
| STEMdiff SMADi Neural Induction Kit | STEMCELL Technologies | 8581 | |
| T3 triiodothyronine | MilliporeSigma | T6397 | |
| Tempo-iOlogo: Human iPSC-derived OPCs | Tempo BioScience | SKU102 | |
| TetO-Ng2-Puro | Addgene | 52047 | |
| VSV-G | Addgene | 12259 |
References
- Pelvig, D. P., Pakkenberg, H., Stark, A. K., Pakkenberg, B. Neocortical glial cell numbers in human brains. Neurobiology of Aging. 29 (11), 1754-1762 (2008).
- Barres, B. A. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 60 (3), 430-440 (2008).
- De Strooper, B., Karran, E. The cellular phase of Alzheimer's disease. Cell. 164 (4), 603-615 (2016).
- Monje, M. Myelin plasticity and nervous system function. Annual Review of Neuroscience. 41, 61-76 (2018).
- Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., Bergles, D. E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nature Neuroscience. 21 (5), 696-706 (2018).
- Gibson, E. M., et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science. 344 (6183), 1252304 (2014).
- Pan, S., Mayoral, S. R., Choi, H. S., Chan, J. R., Kheirbek, M. A. Preservation of a remote fear memory requires new myelin formation. Nature Neuroscience. 23 (4), 487-499 (2020).
- Thornton, M. A., Hughes, E. G. Neuron-oligodendroglia interactions: Activity-dependent regulation of cellular signaling. Neuroscience Letters. 727, 134916 (2020).
- Ettle, B., Schlachetzki, J. C. M., Winkler, J. Oligodendroglia and myelin in neurodegenerative diseases: more than just bystanders. Molecular Neurobiology. 53 (5), 3046-3062 (2016).
- Essayan-Perez, S., Zhou, B., Nabet, A. M., Wernig, M., Huang, Y. A. Modeling Alzheimer's disease with human iPS cells: advancements, lessons, and applications. Neurobiology of Disease. 130, 104503 (2019).
- Li, L., et al. GFAP mutations in astrocytes impair oligodendrocyte progenitor proliferation and myelination in an hiPSC model of Alexander disease. Cell Stem Cell. 23 (2), 239-251 (2018).
- Lin, Y. T., et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer's disease phenotypes in human iPSC-derived brain cell types. Neuron. 98 (6), 1294 (2018).
- TCW, J., et al. Cholesterol and matrisome pathways dysregulated in human APOE ε4 glia. bioRxiv. , (2019).
- Ang, C. E., Wernig, M.
- Penney, J., Ralvenius, W. T., Tsai, L. H. Modeling Alzheimer's disease with iPSC-derived brain cells. Molecular Psychiatry. 25 (1), 148-167 (2020).
- Zhang, Y., et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron. 78 (5), 785-798 (2013).
- Huang, Y. A., Zhou, B., Nabet, A. M., Wernig, M., Sudhof, T. C. Differential signaling mediated by ApoE2, ApoE3, and ApoE4 in human neurons parallels Alzheimer's Disease risk. Journal of Neuroscience. 39 (37), 7408-7427 (2019).
- Huang, Y. A., Zhou, B., Wernig, M., Sudhof, T. C. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Abeta Secretion. Cell. 168 (3), 427-441 (2017).
- Yang, N., et al. Generation of oligodendroglial cells by direct lineage conversion. Nature Biotechnology. 31 (5), 434-439 (2013).
- Douvaras, P., et al. Efficient generation of myelinating oligodendrocytes from primary progressive multiple sclerosis patients by induced pluripotent stem cells. Stem Cell Reports. 3 (2), 250-259 (2014).
- Lee, E. H., Park, C. H. Comparison of reprogramming methods for generation of induced-oligodendrocyte precursor cells. Biomolecules & Therapeutics (Seoul). 25 (4), 362-366 (2017).
- Ehrlich, M., et al. Rapid and efficient generation of oligodendrocytes from human induced pluripotent stem cells using transcription factors. Proceedings of the National Academy of Sciences of the United States of America. 114 (11), 2243-2252 (2017).
- Rodrigues, G. M. C., et al. Defined and scalable differentiation of human oligodendrocyte precursors from pluripotent stem cells in a 3D culture system. Stem Cell Reports. 8 (6), 1770-1783 (2017).
- Hu, B. Y., Du, Z. W., Li, X. J., Ayala, M., Zhang, S. C. Human oligodendrocytes from embryonic stem cells: conserved SHH signaling networks and divergent FGF effects. Development. 136 (9), 1443-1452 (2009).
- Izrael, M., et al. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Molecular and Cellular Neuroscience. 34 (3), 310-323 (2007).
- Yamashita, T., et al. Differentiation of oligodendrocyte progenitor cells from dissociated monolayer and feeder-free cultured pluripotent stem cells. PLoS One. 12 (2), 0171947 (2017).
- Wang, S., et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell. 12 (2), 252-264 (2013).
- Chanoumidou, K., Mozafari, S., Baron-Van Evercooren, A., Kuhlmann, T. Stem cell derived oligodendrocytes to study myelin diseases. Glia. 68 (4), 705-720 (2020).
- Chetty, S., et al. A simple tool to improve pluripotent stem cell differentiation. Nature Methods. 10 (6), 553-556 (2013).
- Li, J., et al. A transient DMSO treatment increases the differentiation potential of human pluripotent stem cells through the Rb family. PLoS One. 13 (12), 0208110 (2018).
- Sambo, D., Li, J., Brickler, T., Chetty, S. Transient treatment of human pluripotent stem cells with DMSO to promote differentiation. Journal of Visualized Experiments: JoVE. (149), (2019).
- Douvaras, P., Fossati, V. Generation and isolation of oligodendrocyte progenitor cells from human pluripotent stem cells. Nature Protocols. 10 (8), 1143-1154 (2015).
- Mei, F., et al. Micropillar arrays as a high-throughput screening platform for therapeutics in multiple sclerosis. Nature Medicine. 20 (8), 954-960 (2014).
- Madhavan, M., et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nature Methods. 15 (9), 700-706 (2018).
- Zhang, Y., et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 89 (1), 37-53 (2016).
- Grubman, A., et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer's disease reveals cell-type-specific gene expression regulation. Nature Neuroscience. 22 (12), 2087-2097 (2019).
- Goldman, S. A., Kuypers, N. J. How to make an oligodendrocyte. Development. 142 (23), 3983-3995 (2015).
- Behrendt, G., et al. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia. 61 (2), 273-286 (2013).
- Patzke, C., et al. Neuromodulator signaling bidirectionally controls vesicle numbers in human synapses. Cell. 179 (2), 498-513 (2019).
- Piao, J., et al. Human embryonic stem cell-derived oligodendrocyte progenitors remyelinate the brain and rescue behavioral deficits following radiation. Cell Stem Cell. 16 (2), 198-210 (2015).
- Keirstead, H. S., et al. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. Journal of Neuroscience. 25 (19), 4694-4705 (2005).
- Kim, D. S., et al. Rapid generation of OPC-like cells from human pluripotent stem cells for treating spinal cord injury. Experimental & Molecular Medicine. 49 (7), 361 (2017).
Tags
Nörobilim Sayı 165 indüklenmiş pluripotent kök hücreler iPS hücreleri insan embriyonik kök hücreleri ES hücreleri nöronlar oligodendrositler oligodendrosit progenitör hücreleri OPC'ler Alzheimer hastalığı amiloid-beta peptitleri sinapslarErratum
Formal Correction: Erratum: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions
Posted by JoVE Editors on 12/29/2020.
Citeable Link.
An erratum was issued for: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions. The Representative Results section has been updated.
Figure 3 was updated from:
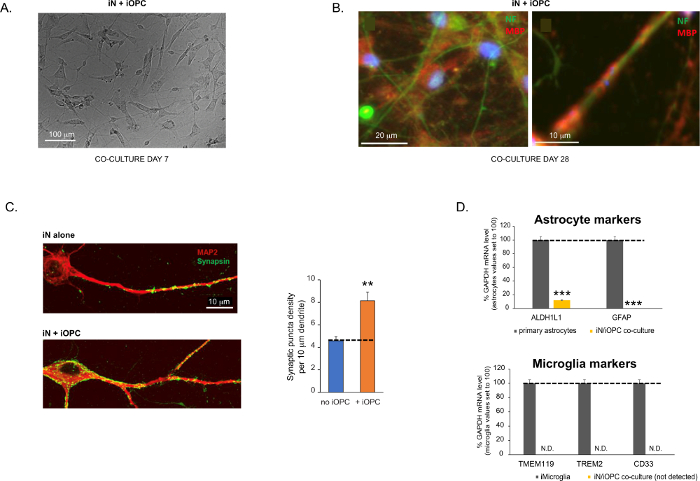
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. Please click here to view a larger version of this figure.
to:
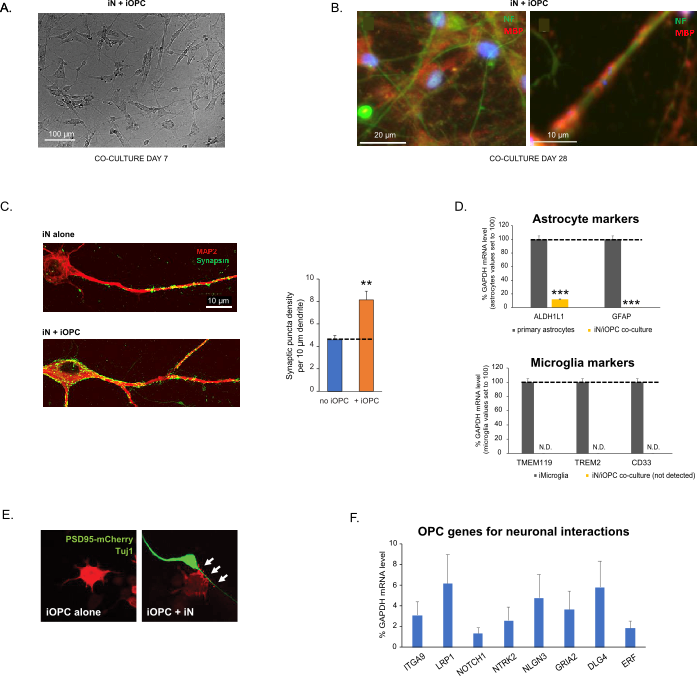
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. (E) Coculturing iOPC with iN leads to the formation of neuron-OPC synapses. The fluorescence-tagged post-synaptic marker PSD95-mCherry is expressed only in OPCs, and display a diffuse pattern in single cultures (left) but aggregate to form puncta in cocultures (right, indicated by arrows; Tuj1, neuronal marker). (F) The expression of well-characterized oligodendroglial genes that can sense and respond to neuronal activities in the pure cultures of iOPCs at Day 14. Please click here to view a larger version of this figure.
The fourth paragraph was updated from:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D).
to:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D). The iOPCs express a good number of well-characterized genes that can potentially respond to and mediate the activity-dependent signals from neighboring neurons, in a paracrine (e.g. neurotrophins and metabolites) and/or a synaptic manner (Figure 3E and 3F).
