


ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Die Neuron-Glia-Interaktionen bei der Neurodegeneration sind aufgrund unzureichender Werkzeuge und Methoden nicht gut verstanden. Hier beschreiben wir optimierte Protokolle zur Gewinnung induzierter Neuronen, Oligodendrozytenvorläuferzellen und Oligodendrozyten aus humanen pluripotenten Stammzellen und geben Beispiele für die Werte dieser Methoden zum Verständnis zelltypspezifischer Beiträge bei der Alzheimer-Krankheit.
Abstract
Bei der Alzheimer-Krankheit (AD) und anderen neurodegenerativen Erkrankungen ist das oligodendrogliale Versagen ein häufiges frühes pathologisches Merkmal, aber wie es zur Entwicklung und Progression der Krankheit beiträgt, insbesondere in der grauen Substanz des Gehirns, bleibt weitgehend unbekannt. Die Dysfunktion von Oligodendrozyten-Linienzellen ist gekennzeichnet durch Mängel in der Myelinisierung und eine gestörte Selbsterneuerung von Oligodendrozyten-Vorläuferzellen (OPCs). Diese beiden Defekte werden zumindest teilweise durch die Störung der Interaktionen zwischen Neuronen und Oligodendrozyten während des Aufbaus der Pathologie verursacht. OPCs führen während der ZNS-Entwicklung zu myelinisierenden Oligodendrozyten. In der reifen Hirnrinde sind OPCs die wichtigsten proliferativen Zellen (die ~5% der gesamten Gehirnzellen ausmachen) und kontrollieren die Neubildung von Myelin in Abhängigkeit von der neuronalen Aktivität. Solche Neuron-zu-Oligodendrozyten-Kommunikationen sind insbesondere im Zusammenhang mit neurodegenerativen Erkrankungen wie AD aufgrund des Mangels an geeigneten Werkzeugen deutlich unteruntersucht. In den letzten Jahren haben unsere Gruppe und andere bedeutende Fortschritte bei der Verbesserung der derzeit verfügbaren Protokolle zur individuellen Erzeugung funktioneller Neuronen und Oligodendrozyten aus menschlichen pluripotenten Stammzellen gemacht. In diesem Manuskript beschreiben wir unsere optimierten Verfahren, einschließlich der Etablierung eines Kokultursystems zur Modellierung der Neuron-Oligodendrozyten-Verbindungen. Unsere illustrativen Ergebnisse deuten auf einen unerwarteten Beitrag von OPCs / Oligodendrozyten zur Amyloidose und Synapsenintegrität des Gehirns hin und unterstreichen den Nutzen dieser Methodik für die AD-Forschung. Dieser reduktionistische Ansatz ist ein mächtiges Werkzeug, um die spezifischen heterozellulären Interaktionen aus der inhärenten Komplexität im Gehirn zu sezieren. Es wird erwartet, dass die hier beschriebenen Protokolle zukünftige Studien zu oligodendroglialen Defekten in der Pathogenese der Neurodegeneration erleichtern.
Introduction
Oligodendrozyten-Linienzellen – einschließlich Oligodendrozyten-Vorläuferzellen (OPCs), myelinisierende Oligodendrozyten und Übergangstypen dazwischen – bilden eine Hauptgruppe menschlicher Gehirnzellen1, die aktiv an vielen kritischen Funktionen für den ordnungsgemäßen Betrieb und die Aufrechterhaltung unseres zentralen Nervensystems während der neuronalen Entwicklung und des Alterns beteiligtsind 2,3,4 . Während Oligodendrozyten dafür bekannt sind, Myelin zu produzieren, um die Übertragung neuronaler Aktivität zu erleichtern und die axonale Gesundheit in der weißen Substanz zu unterstützen, sind OPCs reichlich vorhanden (~ 5%) in der grauen Substanz, wo Myelinisierung knapp ist und aktivitätsabhängige Signalfunktionen ausführen, um das Lernverhalten und die Gedächtnisbildung zu steuern 5,6,7,8 . Wie oligodendrogliale Zellen funktionieren und dysfunktional in der Pathogenese der Alzheimer-Krankheit (AD) und anderer altersassoziierter neurodegenerativer Erkrankungen ist zu wenig untersucht9. Die Unzulänglichkeiten eines geeigneten Modellsystems und Defizite im Allgemeinwissen, um einen experimentellen Weg nach vorne zu leiten, sind die Hauptgründe für diese Lücke.
Angesichts der jüngsten Durchbrüche bei der Gewinnung menschlicher Gehirnzellen aus pluripotenten Stammzellen, einschließlich embryonaler Stammzellen (ES) und induzierter pluripotenter Stammzellen (iPS), haben sich solche zellulären Modelle in Verbindung mit modernen Gen-Editing-Tools als robuste Werkzeuge zur Handhabung des komplizierten Nexus zellulärer Interaktionen im Gehirn erwiesen und sind in der Lage, humanspezifische Krankheitsmanifestationennachzuweisen10. 11. In Anbetracht der Tatsache, dass einzelne Gehirnzelltypen angesichts der gleichen AD-fördernden Bedingungen unterschiedliche und sogar widersprüchliche Wirkungen zeigen können12,13, bietet diese Stammzellmethodik auf einzigartige Weise zelltypspezifische Informationen, die zuvor unter Verwendung etablierter In-vivo- oder In-vitro-Modelle, die nur aggregierte Messwerte aus Sammlungen von Gehirnzelltypen liefern, übersehen wurden. In den letzten zehn Jahren wurde eine große Anzahl zuverlässiger Protokolle entwickelt, um menschliche Neuronen aus Transdifferenzierung von ES/iPS-Zellen oder direkter Umwandlung aus anderen terminaldifferenzierten Zelltypen (z. B. Fibroblasten) zu erzeugen14,15. Insbesondere die Anwendung wichtiger neurogener Transkriptionsfaktoren (z.B. Neurogenin 2, Ngn2)16 auf humane pluripotente Stammzellen kann eine homogene Population gut charakterisierter neuronaler Zelltypen für Reinkulturen erzeugen, ohne dass eine Kokulturierung mit Gliazellen erforderlich ist12,17,18. Für induzierte menschliche Oligodendrozyten gibt es einige veröffentlichte Protokolle, die funktionelle Zellen erzeugen können, die ihren primären Gegenstücken sehr ähnlich sind, mit einer breiten Palette von Effizienz und Bedarf an Zeit und Ressourcen 19,20,21,22,23,24,25,26,27,28 . Bisher wurde keines dieser Protokolle angewendet, um zu untersuchen, wie oligodendrogliale Zellen auf die AD-Pathogenese reagieren und diese beeinflussen.
Hier beschreiben wir unsere verbesserten Protokolle für Einzel- und Mischkulturen von humaninduzierten Neuronen (iNs) und OPCs/Oligodendrozyten (iOPCs/iOLs). Das hier beschriebene iN-Protokoll basiert auf dem weit verbreiteten Ngn2-Ansatz16 und hat zusätzlich die Eigenschaft, gliafrei zu sein. Die resultierenden iNs sind homogen und ähneln stark den exzitatorischen Neuronen der kortikalen Schicht 2/3 mit charakteristischer pyramidaler Morphologie, Genexpressionsmuster und elektrophysiologischen Merkmalen17,18 (Abbildung 1). Um einige der grundlegenden Barrieren bei der gerichteten Differenzierung pluripotenter Stammzellen zu überwinden, haben wir eine einfache und effektive Methode der Vorbehandlung von niedrig dosiertem Dimethylsulfoxid (DMSO) entwickelt29,30 und berichteten über eine erhöhte Neigung menschlicher ES/iPS-Zellen, in iOPCs und iOLs31 zu transdifferenzieren, basierend auf einem weitgehend angepassten Protokoll von Douvaras und Fossati 32 . Wir haben das Protokoll weiter vereinfacht und eine robuste differenzierungsfördernde Verbindung, Clemastin 7,33,34, integriert, um den Prozess der oligodendroglialen Reifung zu beschleunigen. Als Ergebnis (Abbildung 2) können die iOPCs in 2 Wochen (~95% positiv für den Marker O4) und iOLs in vier Wochen (exprimierend für reife Marker MBP und PLP1) generiert werden. Interessanterweise fanden wir heraus, dass iOPCs allein eine bemerkenswerte Menge an Amyloid-β (Aβ) sezernieren, was mit den unabhängigen transkriptomischen Daten übereinstimmt, die die reichliche Expression des Amyloid-Vorläuferproteins (APP) und der Verarbeitungsprotease β-Sekretase (BACE1) in Oligodendrozyten-Linienzellen35,36 zeigen. Darüber hinaus fördert unser iN-iOPC-Kokultursystem die Umhüllung von Axonen durch MBP-positive iOL-Prozesse und bietet eine signifikante Unterstützung für die Synapsenbildung (Abbildung 3). Daher haben die Protokolle, die wir unten beschrieben haben, technische und biologische Vorteile gegenüber zuvor katalogisierten Neuron-Oligodendroglia-Co-Kultivierungsmethoden und versprechen eine bessere Modellierung der Neurodegeneration bei AD.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Induktion menschlicher Neuronen aus humanen pluripotenten Stammzellen
- Lentivirus-Vorbereitung (~5 Tage, detailliertes Protokoll wie zuvor beschrieben16)
- Platte ~ 1 Million HEK293T-Zellen jeder T75-Kolben, um sie ~ 40% konfluent zu haben, wenn Transfektion durchgeführt wird. Transfizieren Sie sie mit Plasmiden, die Tetracyclin-induzierbares Ngn2 und puromycin-resistentes Gen (PuroR; unter derselben TetO-Promotorkontrolle), rtTA und die drei Helferplasmide pRSV-REV, pMDLg/pRRE und VSV-G (12 μg lentivirale Vektor-DNA und 6 μg jeder der Helferplasmid-DNA) exprimieren. Bereiten Sie mindestens drei Flaschen pro Lentivirus-Präparat vor. Verwenden Sie PEI für die Transfektion gemäß den Anweisungen des Herstellers. Wechseln Sie die Medien nach 16 h und verwerfen Sie sie.
- Ernte freigesetzte Viruspartikel durch tägliches Sammeln von Kulturmedien und Ersetzen durch frische Medien für 3 Tage. Bündeln Sie die gesammelten Medien mit viralen Partikeln zur Reinigung. Filtern Sie das Virus durch einen 0,22 μm Filter und zentrifugieren Sie es bei 49.000 x g für 90 min. Resuspendieren Sie das Pellet in der entsprechenden Menge PBS-Glucose (~150 μL).
- Neuroneninduktion (~5 Tage)
HINWEIS: Dieses Induktionsprotokoll (Abbildung 1A; Flussdiagramm) ist sowohl für iPS- als auch für ES-Zellen mit validierter Pluripotenz (die durch immunhistochemische Färbung gut charakterisierter Pluripotenzmarker untersucht werden kann; Abbildung 1B).- Verwenden Sie kommerziell erhältliche menschliche H1-ES-Zellen am Durchgang von 52 (siehe Materialtabelle). Kulturieren Sie die Zellen auf extrazellulärer Matrixlösung, beschichtet mit 6-Well-Platten (~0,5 mg Matrixlösung pro 6-Well-Platte; siehe Materialtabelle) unter Verwendung von ES-Zellerhaltungsmedium (siehe Materialtabelle) und inkubieren Sie die Platten bei 37 °C mit 5%CO2.
- An Tag -2 ES-Zellen (80% konfluent) mit 1 ml Zellablösungslösung (siehe Materialtabelle) ablösen und bei Raumtemperatur 10 min inkubieren. Übertragen Sie die Zellen in eine Röhre; Waschen Sie den Brunnen mit 2 ml Medien und mischen Sie ihn im selben Röhrchen. Zentrifugieren bei 300 x g für 5 min, resuspendieren Sie das Pellet in Medien und platen Sie die Zellen auf matrixbeschichtete 6-Well-Platten mit der Aussaatdichte von 1 x 105 Zellen pro Well.
- An Tag -1 werden Lentiviren, die Ngn2 plus PuroR und rtTA exprimieren, zusammen mit Polybren (8 μg/ml) in die ES-Zellen in frischem ES-Zellerhaltungsmedium gegeben (siehe Materialtabelle). Die genaue Menge an Viren sollte durch tatsächliche Titer oder die Titration bestimmt werden. Wir fügen typischerweise 5 μL jedes Virus pro Vertiefung in eine 6-Well-Platte hinzu.
- An Tag 0 Doxycyclin (2 μg/ml, um die Ngn2-Expression zu aktivieren) in DMEM-F12-Medium mit N2-Ergänzung ohne Morphogene hinzufügen.
- An Tag 1 fügen Sie Puromycin in frischem Medium aus DMEM-F12 plus N2 und Doxycyclin auf die Endkonzentration von 1 μg / ml Medium hinzu. Wählen Sie die transduzierten Zellen in Puromycin für mindestens 24 h aus. Eine höhere Puromycin-Konzentration (bis zu 5 μg/ml) und ein längerer Selektionszeitraum (bis zu 48 h) können erforderlich sein, um die untertransduzierten Zellen angemessen zu entfernen, wenn der Virustiter niedrig ist.
- An Tag 2 trennen Sie differenzierende Neuronen mit Zellablösungslösung (siehe Materialtabelle) und beschichten Sie sie auf 24-Well-Platten (zwischen 80.000 und 200.000 Zellen / Well), die mit Matrixlösung beschichtet sind (siehe Tabelle der Materialien), und halten Sie sie in NBA / B27-Medium ohne Doxycyclin. Die Aussaatdichte ist entscheidend.
- In diesem Stadium können abgelöste Neuronen in einem speziellen kommerziellen Gefriermedium eingefroren werden (siehe Materialtabelle) und bis zu 3 Monate in flüssigem Stickstoff gelagert werden. Reine Neuronen können unter Berücksichtigung des typischen ~15%-20% Zelltods nach dem Auftauen plattiert, allein kultiviert oder mit anderen Gehirnzelltypen co-kultiviert werden (siehe Schritt 3.2.3. für Co-Kultivierung mit OPCs).
- Kultur reiner iNs auf den Platten, die mit extrazellulären Lösungen auf Matrixbasis beschichtet sind, gemäß den Anweisungen des Herstellers (siehe Materialtabelle). Die charakteristische pyramidale Morphologie sollte bis Tag 4 (und Tag 6; Abbildung 1C). Die Synapsenbildung kann bereits am Tag 14 bis 16 nachgewiesen werden und ist am Tag 24 durch immunhistochemische Färbung mit Standard-prä- und postsynaptischen Markern prominent. (Abbildung 1D; beschriftet mit dem präsynaptischen Marker Synapsin 1 und dem dendritischen Marker Map2).
2. Induktion humaner Oligodendrozyten-Vorläuferzellen (OPCs) aus pluripotenten Stammzellen und Oligodendrozytenreifung
- Neuronale Vorläuferzelle (NPC) Generation: Monolayer-Protokoll (~7 Tage). Das Flussdiagramm ist in Abbildung 2A dargestellt.
- Kultur menschlicher H1-ES-Zellen wie zuvor beschrieben (siehe Schritt 1.2.1.) und transdifferenzieren Sie sie in neuronale Vorläuferzellen (NPCs) durch einen etablierten Ansatz namens duale SMADi mit niedermolekularen Inhibitoren für mehrere Signalwege. Hier verwenden wir ein weithin akzeptiertes kommerzielles Kit und folgen dem vom Hersteller bereitgestellten Monolayer-Protokoll (siehe Materialtabelle).
- An Tag -1, Platte 0,5–1 x 10 6 Zellen pro Vertiefung in einer 6-Well-Platte, beschichtet mit einer wachstumsfaktorreduzierten Matrixlösung (siehe Tabelle der Materialien; ~0,5 mg Matrixlösung pro 6-Well-Platte) mit ES-Zellerhaltungsmedium (siehe Tabelle der Materialien). Diese wachstumsfaktorreduzierte Matrixlösung wird verwendet, um alle Platten zu beschichten, die in den folgenden Schritten verwendet werden.
- Am Tag 0 behandeln Sie die Zellen für 24 Stunden mit ES-Zellerhaltungsmedium (siehe Materialtabelle), ergänzt durch 2% DMSO.
- Wechseln Sie an Tag 1–6 das gesamte Medium mit einem warmen (37 °C) neuralen Induktionsmedium, das die SMAD-Inhibitoren aus dem kommerziellen Kit enthält (siehe Materialtabelle). Wenn sich Zellen vor Tag 7 teilen und den Zusammenfluss erreichen, gehen sie auf die Seeddichte von 0,5–1 x 106, wie weiter oben in Schritt 2.1.2 beschrieben.
- An Tag 7 werden NPCs mit Zellablösungslösung (siehe Materialtabelle) und Platte bei einer Aussaatdichte von 1–2 x 105 Zellen/Vertiefung einer 24-Well-Platte durchgehen.
- Bestimmen Sie die Differenzierungseffizienz durch immunhistochemische (IHC) Färbung auf Abwesenheit von Pluripotenzmarker, OCT4 zum Beispiel, und Vorhandensein von NPC-Markern wie PAX6, Nestin und Sox1.
- In diesem Stadium können abgelöste NPCs in den spezialisierten kommerziellen NPC-Gefriermedien eingefroren werden (siehe Materialtabelle) und bis zu 3 Monate in flüssigem Stickstoff gelagert werden. Nach dem Einfrieren und Auftauen für einmal behalten NPCs immer noch die Multipotenz, um Neuronen, Astrozyten und OPCs mit zuverlässigen Protokollen hervorzubringen.
- Bildung von Oligodendrozytenvorläuferzellen (OPC) (~7 Tage). Das Flussdiagramm finden Sie in Abbildung 2A .
- An Tag 7 werden NPCs mit Zellablösungslösung durchgelassen (siehe Materialtabelle) und sie mit einer Seeding-Dichte von 1–2 x 105 Zellen pro Vertiefung in eine 24-Well-Platte in warmem (37 °C) neuronalem Induktionsmedium plus SMAD-Inhibitoren aus dem kommerziellen Kit (siehe Materialtabelle) geblasen.
- An Tag 8 eine Lösung von 1% DMSO im OPC-Differenzierungsmedium vorbereiten und die plattierten NPCs 24 h lang behandeln. Das OPC-Differenzierungsmedium besteht aus: DMEM/F12-Medium, 1% N2-Ergänzung, 1% B27-Ergänzung, bFGF bei 20 ng/ml, SAG bei 1 μM, PDGF-AA bei 10 ng/ml (siehe Materialtabelle).
- Ersetzen Sie an Tag 9 die Medien durch ein frisches OPC-Differenzierungsmedium ohne DMSO. Füttern Sie die Zellen jeden zweiten Tag bis zum 15. Tag. Wenn die Zellen vor Tag 15 den Zusammenfluss erreichen, werden sie auf die Aussaatdichte von 1–2 x 105 Zellen pro Vertiefung geleitet, wie in Schritt 2.2.1 beschrieben.
- An Tag 14 werden OPCs in OPC-Differenzierungsmedium mit einer Dichte von 1–2 x 105 Zellen/Vertiefung in einer 24-Well-Platte verkleidet.
- In diesem Stadium (Tag 15) Testzellen auf das Vorhandensein von OPC-spezifischen Markern durch IHC-Färbung oder qPCR (z. B. O4, Olig1/2, CSPG4/Ng2, NKX2.2, PDGFRa; Abbildung 2B) und für das Fehlen von NPC-Markern (Pax6 oder Nestin; Abbildung 2D). Wir weisen typischerweise die O4-Immunreaktivität in mehr als 95% der Zellen am Tag 15 nach. Von besonderer Bedeutung für die Alzheimer-Krankheit ist die Expression von APP (Amyloid-Vorläuferprotein), BACE1 (die Verarbeitungsprotease β-Sekreatase 1) und Peptid-Amyloid-β (Aβ) in OPCs reichlich vorhanden (Abbildung 2F).
- Oligodendrozyten (OL) Reifung (~7–20 Tage)
- Ersetzen Sie an Tag 15 die Medien durch ein OL-Reifungsmedium: Neurobasal-A-Medium, 2% B27-Ergänzung, 1 μM cAMP, 200 ng/ml T3-Trijodthyronin und Clemastin von 1 μM (siehe Materialtabelle). Wechseln Sie das Medium bei Bedarf jeden zweiten Tag oder jeden Tag.
- Wenn die Zellen einen Zusammenfluss von 90% erreichen, teilen Sie sich im Verhältnis 1:3 bis zu 2 Passagen oder bis sich die Zellteilung erheblich verlangsamt. Wenn sich OPCs zu schnell teilen und in weniger als 3 Tagen Konfluenz erreichen, fügen Sie Ara-C (siehe Materialtabelle) in einer Konzentration von 2-5 μM für 1-3 Tage hinzu. Aktive Proliferation deutet auf eine verminderte Reifungseffizienz hin.
- Untersuchen Sie die Effizienz der oligodendroglialen Reifung durch Beurteilung der Expression von OL-Markern, z. B. CLDN11, PLP1, MBP durch qPCR, IHC-Färbung oder Immunoblotting. Die charakteristische Morphologie hochkomplexer Strukturen (Abbildung 2C) und die Expression von OL-Markern (Abbildung 2E) sollten bis Tag 28 leicht nachgewiesen werden.
3. Co-Kultivierung von humanen induzierten Neuronen (iNs) und Oligodendrozyten-Vorläuferzellen (iOPCs)
- iOPC-Beschichtung (~3 Tage)
- Platten-iOPCs am Tag 14 mit einer Dichte von 1 x 105 Zellen pro Vertiefung in einer 24-Well-Platte (wie oben in Schritt 2.2.4 beschrieben) in OPC-Differenzierungsmedium (wie in Schritt 2.2.2 beschrieben).
- iN-iOPC Co-Kultur eingerichtet
- Am Tag 15 lösen Sie die induzierten menschlichen Neuronen im Schritt von Tag 2 nach der Puromycin-Selektion (wie in Schritt 1.2.6 beschrieben) mit Zellablösungslösung ab (siehe Materialtabelle).
- Fügen Sie Neuronen auf die kultivierten OPCs hinzu und plattieren Sie mit der Seeding-Dichte von 2 x 105 Zellen pro Vertiefung in der 24-Well-Platte mit wachsenden OPCs (aus Schritt 3.1.1). Verwenden Sie das Kokulturmedium, das Neurobasal-A-Medium, 2% B27-Ergänzung und 100 ng / ml T3-Trijodthyronin enthält. Wechseln Sie das Medium am nächsten Tag und danach jeden zweiten Tag. Wenn sich OPCs zu schnell vermehren und in weniger als 3 Tagen Konfluenz erreichen, fügen Sie Ara-C in einer Konzentration von 2-5 μM hinzu. Ein repräsentatives Bild der iNs und iOPCs, die nach 7 Tagen in Kokultur gezüchtet wurden, ist in Abbildung 3A dargestellt.
- Verwenden Sie gefrorene Neuronen, die wie oben in Schritt 1.2.7 beschrieben für die Co-Kultivierung mit OPCs vorbereitet wurden. Plattengefrieren und Auftauen von Neuronen mit einer höheren Dichte von 3 x 105 Zellen pro Well.
- Nach Tag 14-16 in Kokulturen kann die Synapsenbildung in iNs durch IHC-Färbung von prä- und postsynaptischen Markern beobachtet werden, und bis Tag 21 sollte die synaptische Puncta reichlich vorhanden sein (Abbildung 3C) und neuronale Aktivitäten können zuverlässig aufgezeichnet werden.
- Ab Tag 21 Testzellen auf OL-spezifische Marker (z. B. MBP und PLP1). Am 28. Tag beobachten wir normalerweise das Phänomen der Umhüllung von iN-Axonen durch iOL-Prozesse, markiert durch IHC-Färbung für spezifische Marker (Abbildung 3B; Neurofilament-NF für iN-Axone und MBP für iOPC-Prozesse).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Direkte Erzeugung von humaninduzierten Neuronen aus humanen pluripotenten Stammzellen
Es ist sehr wichtig, dass die startenden humanen pluripotenten Stammzellen einen hohen Grad an Pluripotenz für die erfolgreiche Erzeugung von iNs oder iOPCs/iOLs aufweisen. Daher sollten Zellen auf bestimmte Marker wie Oct4 und SOX2 gefärbt werden, bevor eines der im vorliegenden Manuskript beschriebenen Induktionsprotokolle gestartet wird (Abbildung 1A). Menschliche H1-Zellen wurden verwendet, um induzierte exzitatorische Vorderhirnneuronen nach dem zuvor veröffentlichten Protokoll von Zhang et al. mit einigen Modifikationen zu erhalten (Abbildung 1C)12,16,17,18. Hier stellen wir ein Protokoll vor, in dem iNs an Tag 2 in Reinkultur auf Matrixlösung (siehe Materialtabelle) in Abwesenheit jeglicher Feederschicht neu plattiert werden: Gliazellen oder Fibroblasten. Zusätzlich zu den zuvor veröffentlichten Protokollen beobachten wir, dass das Einfrieren von iNs an Tag 2 die Zelllebensfähigkeit nicht signifikant beeinflusst (~15%-20% Zelltod nach dem Auftauen). Reine Neuronen in Kultur beginnen an Tag 14–16 mit der Expression von Synapsin1 (Abbildung 1D). Die Etablierung einer reinen neuronalen Kultur ist sehr wichtig, da bestimmte Faktoren, zum Beispiel der führende AD-Risikofaktor ApoE, von Zellen in der Feederschicht exprimiert werden können und dies die Ergebnisse erheblich verfälschen kann.
iOPC-Generierung und iOL-Reifung wird durch DMSO-Behandlung verbessert
Hier stellen wir ein schnelles und effizientes Protokoll vor, das die Generierung von iOPCs in 2 Wochen und ausgereiften iOLs in 4–5 Wochen ermöglicht (Abbildung 2A). Wir nutzten die zuvor entwickelte Methode der transienten DMSO-Behandlung, um die Differenzierungseffizienz für ES- und iPS-Zellen zu erhöhen 29,30,31. Die DMSO-Behandlung bereichert die Anzahl der Zellen in der frühen G1-Phase für eine bessere Signalintegration und begünstigt die Differenzierung. Wir führten die erste Behandlung durch, bevor wir menschliche ES-Zellen dazu brachten, NPCs zu erzeugen, und die zweite Behandlung, bevor wir NPCs in iOPCs differenzierten. Wir können spezifische OPC-Marker (Olig2, CSPG4, NKX2.2 und PDGFRA) bereits 2 Wochen nach der Beschichtung von ES-H1-Zellen nachweisen (Abbildung 2B,E). Die iOPC-Population in diesem Stadium ist ziemlich homogen, mit >95% der Zellen, die positiv für die O4-Färbung sind, und einem hohen Maß an Immunreaktivität für andere Marker (Abbildung 2B). Nach Beginn der OL-Reifung am Tag 15 können wir typischerweise spezifische OL-Marker (MBP, O1, CLDN11 und PLP1) ab Tag 28 erkennen (Abbildung 2C,E). Die Expression dieser stadienspezifischen Marker korreliert mit dem Entwicklungsverlauf von Oligodendrogliazellen und deutet auf ein beschleunigtes Tempo hin, wobei die NPC-Marker progressiv sinken, die OPC-Marker ihren Höhepunkt in der zweiten Woche erreichen und die OL-Marker in der dritten Woche ansteigen (Abbildung 2D,E)37. Bitte beachten Sie, dass dieser Reifungsprozess die Zellpopulationen diversifiziert. Die Subpopulationen im Kontinuum, die mehrere Zwischenstufen zwischen OPCs und reifen myelinisierenden Oligodendrozyten umfassen, können vorhanden sein und einen unterschiedlichen Prozentsatz der Gesamtzellen ausmachen, wobei reifere Zellen zu einem späteren Zeitpunkt dominieren.
Zum Vergleich: Wir haben die hochreferenzierten iOPCs gekauft und nach Herstellerprotokoll in iOL ausgereift. Wir haben die Expression der oben genannten Marker sowohl in unseren iOPC- als auch in unseren iOL-Präparaten und in den von uns gekauften Zellen getestet. Wir stellten fest, dass die nach unserem Protokoll erzeugten Zellen eine höhere Expression aller getesteten Gene aufwiesen (Abbildung 2E). Als wir die sezernierten Konzentrationen von zwei Hauptisoformen von Amyloid-β (Aβ40 und Aβ42) in iNs im Vergleich zu iOPCs testeten, stellten wir interessanterweise fest, dass iOPCs mehr von beiden Fragmenten sezernierten, aber das Verhältnis blieb gleich (Abbildung 2F).
Co-Kultivierung von iNs und iOPCs
Dieses Protokoll ist speziell für die Co-Kultivierung von iNs und iOPCs optimiert und ermöglicht eine Echtzeitüberwachung der interzellulären Kommunikation zwischen diesen beiden Zelltypen im Verlauf der neuronalen Entwicklung. Die idealen Beschichtungsdichten für beide Zelltypen müssen mit einer Reihe von Zellzahltitrationen bestimmt werden, um eine korrekte Differenzierung zu erreichen (Abbildung 3A). Nach 4 Wochen in Kokulturen wird erwartet, dass die iOPCs ausreichend in OLs differenziert werden, die für bestimmte Marker wie MBP positiv sind und Prozesse auf umhüllende Axone ausdehnen (Abbildung 3B). Das Kokultursystem kann die Anzahl der Synapsen robust erhöhen, was darauf hindeutet, dass die iOPCs eine neuronale Unterstützung durch physische Kontakte oder die Freisetzung trophischer Faktoren bieten (Abbildung 3C). Wir können die Kokulturen bis zu 6 Wochen in akzeptabler Gesundheitszustand halten und beobachten, dass die Synapsenzahl und andere neuronale Attribute um die fünfte Woche herum ein Plateau erreichen. Bemerkenswert ist, dass Astrozyten und Mikroglia in unseren Präparaten nicht vorhanden sind und ihre Abwesenheit durch Überprüfung der Expression spezifischer Marker dokumentiert werden kann (Abbildung 3D). Die iOPCs exprimieren eine gute Anzahl gut charakterisierter Gene, die möglicherweise auf die aktivitätsabhängigen Signale benachbarter Neuronen reagieren und diese vermitteln können, und zwar parakrin (z.B. Neurotrophine und Metaboliten) und/oder synaptisch (Abbildung 3E und 3F).

Abbildung 1: Direkte Erzeugung von humaninduzierten Neuronen (iNs) aus hPSCs. (A) Flussdiagramm der iN-Erzeugung. (B) Repräsentative Hellfeld- und Immunfluoreszenzbilder der Ausgangskultur humaner pluripotenter Stammzellen (H1) zur Bestätigung der Pluripotenz. Oct4 wird rot und Sox2 grün angezeigt. (C) Repräsentative Hellfeldbilder von iNs an Tag 4 und Tag 6. (D) Die charakteristische Morphologie für dendritische Arborisation und Synapsenpuncta in iNs, die 24 Tage lang in Reinkultur gezüchtet und durch Immunfluoreszenzfärbung für dendritische Marker Map2 und präsynaptische Marker Synapsin 1 (Syn1) gefärbt wurden. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.

Abbildung 2: iOPC-Generierung und iOL-Reifung. (A) Flussdiagramm der iOPC- und iOL-Generierung. (B) Repräsentative Hellfeld- und Immunfluoreszenzbilder von iOPCs an Tag 15. Olig2 (Pan-Oligodendroglia-Marker) ist grün, O4 (OPC-Marker) rot und DAPI blau dargestellt. Die Bildgebung ergab, dass >95% der iOPCs positiv für O4 und 25% für Olig2 sind. (C) Repräsentative Hellfeld- und Immunfluoreszenzbilder von iOLs an Tag 28. MBP wird grün, O1 rot und DAPI blau dargestellt. (D) Die Expression des NPC-Markers PAX6 nimmt in iOPCs an Tag 14 dramatisch ab und sinkt bei OLs an Tag 28 weiter auf Hintergrund, was auf eine robuste NPC-Transdifferenzierung und ein hohes Maß an Homogenität in der iOPC-Population hinweist. (E) Das Zeitverlauf-Expressionsprofil gemeinsamer OPC- und OL-Markergene in Kulturen, die durch das beschriebene Protokoll erzeugt werden, ohne (-DMSO) oder mit (+DMSO) dem Schritt der DMSO-Inkubation (Schritte 2.1.3 und 2.2.2), die zu verschiedenen Zeitpunkten untersucht wurden. Zum Vergleich wurden kommerzielle iOPCs (siehe Materialtabelle) nach Herstellerangaben ausgereift und sowohl iOPCs (iOPC-Tempo) als auch iOLs (iOL-Tempo) auf die gleichen Marker getestet. Wie erwartet, wurde MBP (ein reifer Oligodendrozytenmarker) in den frühen Stadien der Differenzierung in allen getesteten iOPCs nicht nachgewiesen (N.D.). Das DMSO verbesserte die Effizienz der OPC-Differenzierung und OL-Reifung erheblich. (F) Die Produktion und Sekretion von Aβ40 und Aβ42 in reinen iNs- und iOPCs-Kulturen, gemessen mit kommerziellen ELISA-Kits (siehe Materialtabelle) auf Überstand, der aus reinen iNs- und iOPC-Kulturen sowohl am Tag 15 als auch durch Zellzahlen normalisiert wurde (beide bei der Dichte von 200.000 Zellen pro Vertiefung in einer 24-Well-Platte). Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Die Daten in Balkendiagrammen werden als Mittelwert ± REM (Nr. ≥ 3) dargestellt. Die statistische Signifikanz wurde durch den Student's t-Test (*, p < 0,05; ***, p < 0,001) bewertet; in (D) im Vergleich zum NPC; in (E), im Vergleich zur Steuerung iOPC-Tempo; in (F) im Vergleich zu iN.

Abbildung 3: Co-Kultur von iNs und iOPCs. (A) Repräsentatives Hellfeldbild von co-kultivierten iNs und iOPCs an Tag 7, das eine angemessene Dichte für die weitere Reifung zeigt. (B) Repräsentatives Immunfluoreszenzbild von iNs und iOPCs, die 28 Tage lang kokultiviert wurden. Das axonale Marker-Neurofilament NF ist grün und der oligodendrozytäre Marker MBP rot dargestellt. Rechts, ein Segment des iN-Axons, das von einem iOL-Prozess (MBP+) umschlossen wird. (C) Synapsenbildung in 4 Wochen alten Kokulturen. Die Zellen wurden für Synapsin 1 (Syn1, grün) und MAP2 (rot) gefärbt, und synaptische Puncta wurden durch konfokale Analyse der Dichte entlang der dendritischen Segmente quantifiziert, wie beschrieben17,18. (D) In unseren Kokulturen von iNs und iOPCs (7 Tage Co-Kultivierung) ist die Expression der Astrozytenmarker, ALDHL1 und GFAP, minimal (oben), und die Expression der Mikrogliamarker TMEM119, TREM2 und CD33 wird durch qPCR nicht nachgewiesen (N.D.). Die Kontamination durch diese beiden Gliazelltypen ist somit ausgeschlossen. (E) Die Kokulturierung von iOPC mit iN führt zur Bildung von Neuron-OPC-Synapsen. Der fluoreszenzmarkierte postsynaptische Marker PSD95-mCherry wird nur in OPCs exprimiert und zeigt in Einzelkulturen ein diffuses Muster (links), aggregiert jedoch in Kokulturen zu Puncta (rechts, durch Pfeile gekennzeichnet; Tuj1, neuronaler Marker). (F) Die Expression gut charakterisierter oligodendroglialer Gene, die neuronale Aktivitäten in den Reinkulturen von iOPCs am Tag 14 wahrnehmen und darauf reagieren können. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Die Daten in Balkendiagrammen werden als Mittelwert ± REM dargestellt (Nr. ≥ 3). Die statistische Signifikanz wurde durch den Student's t-Test bewertet (**, p < 0,005; ***, p < 0,001); in (C), verglichen mit der Bedingung ohne OPC; in (D), verglichen mit primären Astrozyten im oberen Bild.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Neben der physikalischen und metabolischen Unterstützung zur Stabilisierung der Synapsenstrukturen und zur Erleichterung der salzatorischen Signalleitung durch Myelinisierung können Oligodendrozyten-Linienzellen neuronale Aktivitätsmuster durch schnelle und dynamische Cross-Talks mit Neuronen 5,6,7 formen. Während in der AD-Pathologie die oligodendroglialen Reaktionen zunächst lediglich als sekundär zu Entzündungen und oxidativem Stress angesehen wurden, gibt es nun vielversprechende Beweise, die argumentieren, dass eine beeinträchtigte Myelinintegrität ein frühes pathogenes Ereignis vor dem Auftreten von Aβ-Aggregation und Tau-Hyperphosphorylierung ist9. Darüber hinaus ist die Reparatur der Myelinisierung durch Selbsterneuerung von OPCs imJahr 38 n. Chr. besonders anfällig, ein Prozess, der stark von neuronalen Aktivitäten abhängt. Das Verständnis des Mechanismus zur Unterstützung gesunder Neuron-Oligodendrozyten-Signalwege stellt daher eine hervorragende Gelegenheit dar, neue therapeutische Ziele zu identifizieren.
Das Einzeltranskriptionsfaktor-Ngn2-Protokoll ist eine der am häufigsten verwendeten Techniken zur Erzeugung von Stammzell-abgeleiteten menschlichen Neuronen, und die hier beschriebenen Verfahren sind weitere Verfeinerungen zur Gewinnung reiner neuronaler Kulturen. Unser iOPC/iOL-Protokoll hat eine kürzere Induktionszeit als die zuvor veröffentlichten Studien (4 bis 24 Wochen), mit einer robusten Ausbeute und Reinheit, die mit anderen gängigen Protokollen vergleichbar ist 19,20,22,23,24,25,26,27,28 . Unser Protokoll führt die schrittweise Differenzierung von ES-Zellen zu NPCs, OPCs und schließlich Oligodendrozyten durch charakterisierte Musterhinweise ein und erzeugt funktionelle Zellen, die verwendet werden können, um die Regulation der Myelinisierungshomöostase und Reparatur in vitro oder in vivo (z. B. durch Transplantation in das Shiverer-Mausmodell) zu untersuchen, wie in der vorherigen Arbeit beschrieben. Die Verbesserung unseres Protokolls wird durch die DMSO-Inkubation stark gefördert, die das Retinoblastom-Protein aktiviert und die G1-Phase des Zellzyklus verlängert, um die Reize der gerichteten Differenzierung besser zu integrieren, und auch die terminale Differenzierung in funktionelle Derivateverbessert 29,30. Schließlich verkürzt die Verwendung von Clemastin, einer muskarinischen und antihistaminischen Verbindung, die durch Arzneimittelscreenings für Remyelinisierungstherapeutika33 identifiziert wurde, zusätzlich die Oligodendrozytenreifung, wie sie bei iPS-Zellpräparaten und lebenden Tieren beobachtet wurde 7,28.
Die Einschränkungen der Technik liegen hauptsächlich in der intrinsischen Diskrepanz zwischen den vereinfachten In-vitro-Einstellungen und den In-vivo-Milieus im Gehirn; Diese Diskrepanz führt zu einem Abstrich des vollen Entwicklungspotenzials in den fortgeschrittenen Stadien für einzelne Gehirnzelltypen. Für iNs waren neuere Studien in der Lage, die Kulturen über einen beträchtlich langen Zeitraum in guter synaptischer Gesundheit zu halten, zeigten aber immer noch eine gewisse relative Unreife, die sich in reduzierten wirbelsäulenartigen Strukturen und einer gestörten spontanen synaptischen Übertragung in "alten" iN-Kulturen (selbst den 25 Monate alten) manifestierte39. Während iOPCs reproduzierbar gezeigt haben, dass sie Axone in vivo myelinaten, nachdem sie in transgene Mäusegehirne transplantiert wurden, stellen die In-vitro-Myelinisierungstests mit elektronenmikroskopischer Auswertung für fast alle veröffentlichten Protokolle19,28, einschließlich diesem, immer noch eine technische Herausforderung mit unbefriedigender Effizienz dar. Daher wird nicht erwartet, dass unser Neuron-OPC-Kokultursystem den Alterungsprozess des Gehirns sowie das späte Stadium der AD-Pathologie genau nachahmt. Vielmehr ist es einzigartig in der Lage, die ausgeklügelten interzellulären Interaktionen zwischen Neuronen und OPCs oder Oligodendrozyten im Frühstadium zu entwirren, die unabhängig von Myelinisierung sind und dennoch für die richtige neuronale Entwicklung und Krankheitspathogenese von grundlegender Bedeutung sind.
Jedes der drei hier beschriebenen Verfahren hat seine spezifischen Schritte innerhalb des Protokolls und kann Änderungen und Fehlerbehebung erfordern. Für das iN-Protokoll gibt es zwei kritische Schritte: Puromycin-Auswahl (Schritt 1.2.5) und Plattierungsdichte (Schritt 1.2.6). Die unvollständige Entfernung von untertransduzierten Zellen führt zur Kontamination schlecht differenzierter Zellen und beeinträchtigt das neuronale Überleben und die Funktionen. Die in Schritt 1.2.5 beschriebenen Modifikationen für eine stärkere Puromycin-Selektion mit höherer Konzentration und längerer Inkubation müssten berücksichtigt werden. Die geeignete Beschichtungsdichte sollte durch Titration für jede pluripotente Zelllinie bestimmt werden, da eine geringe Dichte zum Kollaps der Kulturen führt und eine hohe Dichte die Zellaggregation fördert und das neuronale Wachstum behindert. Für das iOPC/iOL-Protokoll sind die beiden kritischen Schritte die Kontrolle der Zellproliferation bei der OPC-Differenzierung (Schritt 2.2.3) und die Plattierungsdichte für die OL-Reifung (Schritte 2.2.4 und 2.3.2). Das Überwachsen differenzierender NPCs signalisiert eine schlechte Reaktion auf die OPC-Differenzierungsreize und muss durch eine geeignete Dosierung der Ara-C-Behandlung (innerhalb des angegebenen Bereichs) gedämpft werden. Bei der Beschichtung von OPCs für die Reifung wird hier ein geringerer Zelldichtebereich bevorzugt, da die spärliche Verteilung die Induktion einer physiologischen Morphologie komplexer Strukturen erleichtern kann (wie in Abbildung 2C gezeigt). Für das iN-iOPC Co-Kultivierungsprotokoll möchten wir auf den kritischen Schritt der Beschichtung mit einer für beide Zelltypen geeigneten Dichte (Schritte 3.1.1 und 3.2.2) hinweisen. Insbesondere binden sich die iNs möglicherweise nicht gut an die Oberfläche zwischen wachsenden OPCs und neigen dazu, sich zuerst zu lösen, wenn die Kultur einen Zusammenfluss erreicht. Das optimale Verhältnis müsste durch Titrieren der Zellzahlen ermittelt werden.
Insgesamt ist dieser reduktionistische Ansatz, der in unseren Protokollen enthalten ist, ein mächtiges Werkzeug, um die spezifischen heterozellulären Interaktionen aus der inhärenten Komplexität des menschlichen Gehirns zu sezieren und dazu zu dienen, die oligodendrogliale Biologie in Gesundheit und AD aufzudecken. Die Bedeutung gegenüber bestehenden Methoden ist daher unserer Meinung nach ziemlich offensichtlich. Ein weiterer Nutzen der hier entwickelten Methoden für die zukünftigen Anwendungen ist die zellbasierte Therapie bei demyelinisierenden Zuständen wie der Strahlenposttherapie40 und der Rückenmarksverletzung41,42. Darüber hinaus kann die Hochdurchsatzkapazität dieses stammzellbasierten Systems auch in größerem Maßstab genutzt werden, um Bibliotheken kleiner Moleküle nach Verbindungen zu durchsuchen, die den physiologischen Status von Neuronen, OPCs, Oligodendrozyten und deren Wechselwirkungen schützen oder wiederherstellen können. Daher glauben wir, dass die hier beschriebenen Protokolle zukünftige Arbeiten zur Entwicklung besserer Modellierungswerkzeuge und wirksamer Behandlungen für AD und andere neurodegenerative Erkrankungen erleichtern werden.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Die Autoren haben nichts offenzulegen.
Acknowledgments
Diese Arbeit wurde durch die Zuschüsse der National Institutes of Health (R00 AG054616 an Y.A.H. und T32 GM136566 an K.C.), der Stanford University School of Medicine und eines Siebel Fellowship (vergeben an S.C.) unterstützt. Y.A.H. ist DaF Translational Professor am Center for Translational Neuroscience am Brown Institute for Translational Sciences.
Materials
| Name | Company | Catalog Number | Comments |
| Accutase | STEMCELL Technologies | 7920 | |
| B27 supplement | ThermoFisher | 17504044 | |
| bFGF | ThermoFisher | PHG 0266 | |
| cAMP | MilliporeSigma | A9501 | |
| Clemastine | MilliporeSigma | SML0445 | |
| DMEM/F12 medium | STEMCELL Technologies | 36254 | |
| DMSO | ThermoFisher | D12345 | |
| Doxycycline | MilliporeSigma | D3072 | |
| Fetal Bovine Serum | ScienCell | 10 | |
| H1 human ES cells | WiCell | WA01 | |
| Matrigel | Corning | 354234 | |
| mTeSR plus | STEMCELL Technologies | 5825 | |
| N2 supplement | ThermoFisher | 17502001 | |
| Neurobasal A medium | ThermoFisher | 10888-022 | |
| Non Essential Amino Acids | ThermoFisher | 11140-050 | |
| PDGF-AA | R&D Systems | 221-AA-010 | |
| PEI | VWR | 71002-812 | |
| pMDLg/pRRE | Addgene | 12251 | |
| Polybrene | MilliporeSigma | TR-1003-G | |
| pRSV-REV | Addgene | 12253 | |
| Puromycin | ThermoFisher | A1113803 | |
| ROCK Inhibitor Y-27632 | STEMCELL Technologies | 72302 | |
| SAG | Tocris | 4366 | |
| STEMdiff Neural Progenitor Freezing Media | STEMCELL Technologies | 5838 | |
| STEMdiff SMADi Neural Induction Kit | STEMCELL Technologies | 8581 | |
| T3 triiodothyronine | MilliporeSigma | T6397 | |
| Tempo-iOlogo: Human iPSC-derived OPCs | Tempo BioScience | SKU102 | |
| TetO-Ng2-Puro | Addgene | 52047 | |
| VSV-G | Addgene | 12259 |
References
- Pelvig, D. P., Pakkenberg, H., Stark, A. K., Pakkenberg, B. Neocortical glial cell numbers in human brains. Neurobiology of Aging. 29 (11), 1754-1762 (2008).
- Barres, B. A. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 60 (3), 430-440 (2008).
- De Strooper, B., Karran, E. The cellular phase of Alzheimer's disease. Cell. 164 (4), 603-615 (2016).
- Monje, M. Myelin plasticity and nervous system function. Annual Review of Neuroscience. 41, 61-76 (2018).
- Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., Bergles, D. E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nature Neuroscience. 21 (5), 696-706 (2018).
- Gibson, E. M., et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science. 344 (6183), 1252304 (2014).
- Pan, S., Mayoral, S. R., Choi, H. S., Chan, J. R., Kheirbek, M. A. Preservation of a remote fear memory requires new myelin formation. Nature Neuroscience. 23 (4), 487-499 (2020).
- Thornton, M. A., Hughes, E. G. Neuron-oligodendroglia interactions: Activity-dependent regulation of cellular signaling. Neuroscience Letters. 727, 134916 (2020).
- Ettle, B., Schlachetzki, J. C. M., Winkler, J. Oligodendroglia and myelin in neurodegenerative diseases: more than just bystanders. Molecular Neurobiology. 53 (5), 3046-3062 (2016).
- Essayan-Perez, S., Zhou, B., Nabet, A. M., Wernig, M., Huang, Y. A. Modeling Alzheimer's disease with human iPS cells: advancements, lessons, and applications. Neurobiology of Disease. 130, 104503 (2019).
- Li, L., et al. GFAP mutations in astrocytes impair oligodendrocyte progenitor proliferation and myelination in an hiPSC model of Alexander disease. Cell Stem Cell. 23 (2), 239-251 (2018).
- Lin, Y. T., et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer's disease phenotypes in human iPSC-derived brain cell types. Neuron. 98 (6), 1294 (2018).
- TCW, J., et al. Cholesterol and matrisome pathways dysregulated in human APOE ε4 glia. bioRxiv. , (2019).
- Ang, C. E., Wernig, M.
- Penney, J., Ralvenius, W. T., Tsai, L. H. Modeling Alzheimer's disease with iPSC-derived brain cells. Molecular Psychiatry. 25 (1), 148-167 (2020).
- Zhang, Y., et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron. 78 (5), 785-798 (2013).
- Huang, Y. A., Zhou, B., Nabet, A. M., Wernig, M., Sudhof, T. C. Differential signaling mediated by ApoE2, ApoE3, and ApoE4 in human neurons parallels Alzheimer's Disease risk. Journal of Neuroscience. 39 (37), 7408-7427 (2019).
- Huang, Y. A., Zhou, B., Wernig, M., Sudhof, T. C. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Abeta Secretion. Cell. 168 (3), 427-441 (2017).
- Yang, N., et al. Generation of oligodendroglial cells by direct lineage conversion. Nature Biotechnology. 31 (5), 434-439 (2013).
- Douvaras, P., et al. Efficient generation of myelinating oligodendrocytes from primary progressive multiple sclerosis patients by induced pluripotent stem cells. Stem Cell Reports. 3 (2), 250-259 (2014).
- Lee, E. H., Park, C. H. Comparison of reprogramming methods for generation of induced-oligodendrocyte precursor cells. Biomolecules & Therapeutics (Seoul). 25 (4), 362-366 (2017).
- Ehrlich, M., et al. Rapid and efficient generation of oligodendrocytes from human induced pluripotent stem cells using transcription factors. Proceedings of the National Academy of Sciences of the United States of America. 114 (11), 2243-2252 (2017).
- Rodrigues, G. M. C., et al. Defined and scalable differentiation of human oligodendrocyte precursors from pluripotent stem cells in a 3D culture system. Stem Cell Reports. 8 (6), 1770-1783 (2017).
- Hu, B. Y., Du, Z. W., Li, X. J., Ayala, M., Zhang, S. C. Human oligodendrocytes from embryonic stem cells: conserved SHH signaling networks and divergent FGF effects. Development. 136 (9), 1443-1452 (2009).
- Izrael, M., et al. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Molecular and Cellular Neuroscience. 34 (3), 310-323 (2007).
- Yamashita, T., et al. Differentiation of oligodendrocyte progenitor cells from dissociated monolayer and feeder-free cultured pluripotent stem cells. PLoS One. 12 (2), 0171947 (2017).
- Wang, S., et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell. 12 (2), 252-264 (2013).
- Chanoumidou, K., Mozafari, S., Baron-Van Evercooren, A., Kuhlmann, T. Stem cell derived oligodendrocytes to study myelin diseases. Glia. 68 (4), 705-720 (2020).
- Chetty, S., et al. A simple tool to improve pluripotent stem cell differentiation. Nature Methods. 10 (6), 553-556 (2013).
- Li, J., et al. A transient DMSO treatment increases the differentiation potential of human pluripotent stem cells through the Rb family. PLoS One. 13 (12), 0208110 (2018).
- Sambo, D., Li, J., Brickler, T., Chetty, S. Transient treatment of human pluripotent stem cells with DMSO to promote differentiation. Journal of Visualized Experiments: JoVE. (149), (2019).
- Douvaras, P., Fossati, V. Generation and isolation of oligodendrocyte progenitor cells from human pluripotent stem cells. Nature Protocols. 10 (8), 1143-1154 (2015).
- Mei, F., et al. Micropillar arrays as a high-throughput screening platform for therapeutics in multiple sclerosis. Nature Medicine. 20 (8), 954-960 (2014).
- Madhavan, M., et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nature Methods. 15 (9), 700-706 (2018).
- Zhang, Y., et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 89 (1), 37-53 (2016).
- Grubman, A., et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer's disease reveals cell-type-specific gene expression regulation. Nature Neuroscience. 22 (12), 2087-2097 (2019).
- Goldman, S. A., Kuypers, N. J. How to make an oligodendrocyte. Development. 142 (23), 3983-3995 (2015).
- Behrendt, G., et al. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia. 61 (2), 273-286 (2013).
- Patzke, C., et al. Neuromodulator signaling bidirectionally controls vesicle numbers in human synapses. Cell. 179 (2), 498-513 (2019).
- Piao, J., et al. Human embryonic stem cell-derived oligodendrocyte progenitors remyelinate the brain and rescue behavioral deficits following radiation. Cell Stem Cell. 16 (2), 198-210 (2015).
- Keirstead, H. S., et al. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. Journal of Neuroscience. 25 (19), 4694-4705 (2005).
- Kim, D. S., et al. Rapid generation of OPC-like cells from human pluripotent stem cells for treating spinal cord injury. Experimental & Molecular Medicine. 49 (7), 361 (2017).
Tags
Neuroscience Ausgabe 165 induzierte pluripotente Stammzellen iPS-Zellen humane embryonale Stammzellen ES-Zellen Neuronen Oligodendrozyten Oligodendrozyten-Vorläuferzellen OPCs Alzheimer-Krankheit Amyloid-beta-Peptide SynapsenErratum
Formal Correction: Erratum: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions
Posted by JoVE Editors on 12/29/2020.
Citeable Link.
An erratum was issued for: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions. The Representative Results section has been updated.
Figure 3 was updated from:
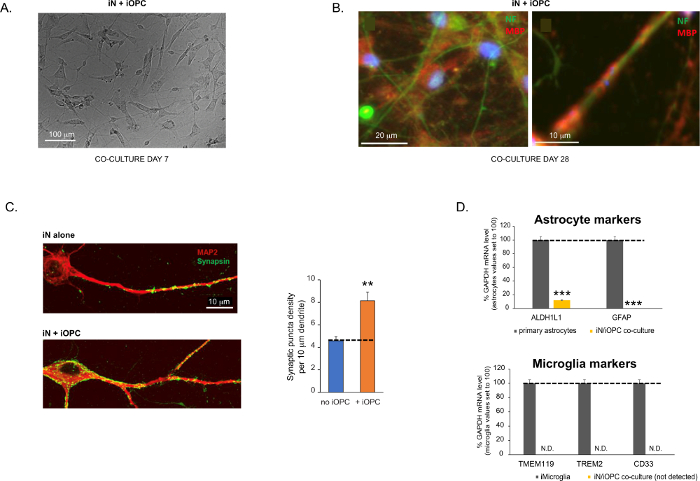
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. Please click here to view a larger version of this figure.
to:
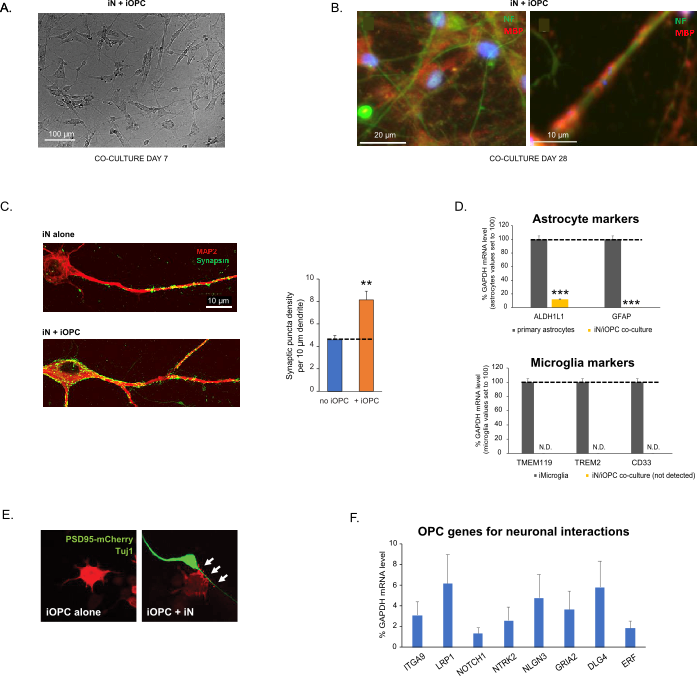
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. (E) Coculturing iOPC with iN leads to the formation of neuron-OPC synapses. The fluorescence-tagged post-synaptic marker PSD95-mCherry is expressed only in OPCs, and display a diffuse pattern in single cultures (left) but aggregate to form puncta in cocultures (right, indicated by arrows; Tuj1, neuronal marker). (F) The expression of well-characterized oligodendroglial genes that can sense and respond to neuronal activities in the pure cultures of iOPCs at Day 14. Please click here to view a larger version of this figure.
The fourth paragraph was updated from:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D).
to:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D). The iOPCs express a good number of well-characterized genes that can potentially respond to and mediate the activity-dependent signals from neighboring neurons, in a paracrine (e.g. neurotrophins and metabolites) and/or a synaptic manner (Figure 3E and 3F).
