Metamerische segmentatie van organismen wordt veel gebruikt in de natuur. Herhaalde structuren zijn essentieel voor de functionaliteit van laterale organen zoals wervels, spieren, zenuwen, bloedvaten, ledematen of bladeren in een lichaamsplan1. Als gevolg van dergelijke fysiologische en geometrische beperkingen van de axiale symmetrie, de meeste phyla van Bilateria-zoals anneliden, geleedpotigen, en chordaten-vertonen segmentatie van hun embryonale weefsels (bijv. ectoderm, mesoderm) antero-posteriorly.
Gewervelde embryo’s segmenteren hun paraxiale mesoderm langs de hoofdas van het lichaam achtereenvolgens in somieten met soortspecifieke intervallen, tellingen en grootteverdelingen. Ondanks deze robuustheid tussen individuele embryo’s binnen een soort, is somietsegmentatie veelzijdig tussen gewervelde soorten. Segmentatie vindt plaats in een uitgebreid regime van tijdsintervallen (van 25 min in zebravissen tot 5 uur bij mensen), maten (van ~20 μm in staart somieten van zebravissen tot ~200 μm in stam somieten van muizen) en tellingen (van 32 in zebravissen tot ~300 in maïsslangen)2. Interessanter is dat visembryo’s zich kunnen ontwikkelen in een breed scala aan temperaturen (van ~ 20,5 °C tot 34 °C voor zebravissen) terwijl ze hun somieten intact houden met de juiste grootteverdelingen door zowel segmentatie-intervallen als axiale reksnelheden te compenseren. Naast dergelijke interessante kenmerken blijft zebravis een nuttig modelorganisme om segmentatie bij gewervelde dieren te bestuderen vanwege de externe, synchrone en transparante ontwikkeling van een plenitude van embryo’s van broers en zussen en hun toegankelijke genetische hulpmiddelen. Vanuit microscopieperspectief ontwikkelen teleostembryo’s zich op een omvangrijke bolvormige dooier, die het gastrulerende weefsel eromheen uitrekt en rondt (figuur 1A). In dit artikel presenteren we een afgeplatte 3D-weefsel explantcultuur voor zebravisstaarten. Dit explantsysteem omzeilt de bolvormige beperkingen van dooiermassa, waardoor toegang wordt verstondkbaar tot live beeldvorming met hoge resolutie van visembryo’s voor somietpatronen.
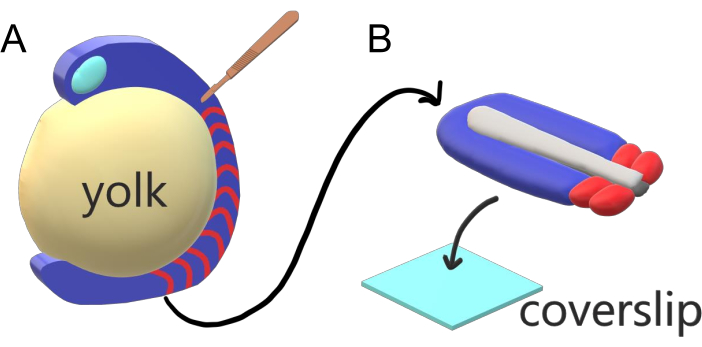
Figuur 1: Slide Chamber Explant System for Zebrafish Embryos. (A) Zebravisembryo’s hebben voordelen voor levende beeldvorming, zoals de transparantie van gastrulerend embryonaal weefsel (blauw), maar het weefsel vormt zich rond een omvangrijke bolvormige dooiermassa (geel) die beeldvorming met een hoge resolutie in intacte embryo’s voorkomt. Staart explants kunnen worden ontleed beginnend met een microchirurgische mes (bruin) gesneden uit het weefsel voorste van somieten (rood) en verder op de grens met de dooier posteriorly. (B) Ontlede staart explants kunnen op een afdeklip (lichtblauw) dorsoventrally worden geplaatst; het houden van neuraal weefsel (lichtgrijs) aan de bovenkant en notochord (donkergrijs) aan de onderkant. Klik hier om een grotere versie van deze afbeelding te bekijken.
