La segmentación metamérica de organismos es ampliamente utilizada en la naturaleza. Las estructuras repetidas son esenciales para la funcionalidad de órganos laterales como vértebras, músculos, nervios, vasos, extremidades u hojas en un plan corporal1. Como resultado de tales restricciones fisiológicas y geométricas de la simetría axial, la mayoría de los filos de Bilateria-tales como anélidos, artrópodos, y cordados-exhiben la segmentación de sus tejidos embrionarios (e.g., ectodermo, mesodermo) antero-posteriorly.
Los embriones de vertebrados segmentan secuencialmente su mesodermo paraxial a lo largo del eje principal del cuerpo en somitas con intervalos específicos de la especie, recuentos y distribuciones de tamaño. A pesar de tal robustez entre embriones individuales dentro de una especie, la segmentación de somita es versátil entre las especies de vertebrados. La segmentación ocurre en un vasto régimen de intervalos de tiempo (de 25 min en el pez cebra a 5 h en humanos), tamaños (de ~20 μm en somitas de cola de pez cebra a ~200 μm en somites de tronco de ratones) y recuentos (de 32 en pez cebra a ~300 en serpientes de maíz)2. Más interesante aún, los embriones de peces pueden desarrollarse en un amplio rango de temperaturas (desde ~ 20.5 ° C hasta 34 ° C para el pez cebra) mientras mantienen sus somitas intactas con distribuciones de tamaño adecuadas al compensar tanto los intervalos de segmentación como las velocidades de elongación axial. Más allá de estas interesantes características, el pez cebra se mantiene como un organismo modelo útil para estudiar la segmentación en vertebrados debido al desarrollo externo, síncrono y transparente de una plenitud de embriones hermanos, así como sus herramientas genéticas accesibles. Negativamente desde una perspectiva de microscopía, los embriones de teleósteo se desarrollan en una yema esférica voluminosa, estirando y redondeando el tejido gastrulante a su alrededor (Figura 1A). En este artículo, presentamos un cultivo de explantes de tejido 3-D aplanado para colas de pez cebra. Este sistema de explantes evita las restricciones esféricas de la masa de yema, lo que permite el acceso a imágenes vivas de alta resolución de embriones de peces para el modelado de somita.
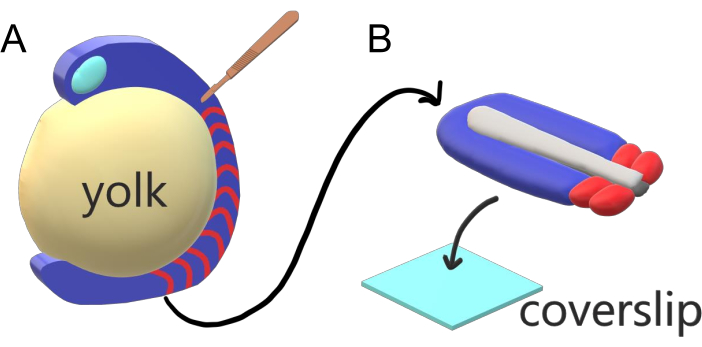
Figura 1:Sistema de explante de cámara deslizante para embriones de pez cebra. (A) Los embriones de pez cebra tienen ventajas para la obtención de imágenes vivas, como la transparencia del tejido embrionario gastrulado (azul), pero el tejido se forma alrededor de una masa de yema esférica voluminosa (amarillo) que impide obtener imágenes casi objetivas y de alta resolución en embriones intactos. Los explantes de cola se pueden diseccionar comenzando con un cuchillo microquirúrgico (marrón) cortado del tejido anterior de somitas (rojo) y continuando en el borde con la yema posterior. (B) Los explantes de cola disecados pueden colocarse en un cubrebocas (azul claro) dorsoventralmente; mantener el tejido neural (gris claro) en la parte superior y la notocorda (gris oscuro) en la parte inferior. Haga clic aquí para ver una versión más amplia de esta figura.
