La segmentation métamérique des organismes est largement utilisée dans la nature. Les structures répétées sont essentielles pour la fonctionnalité des organes latéraux tels que les vertèbres, les muscles, les nerfs, les vaisseaux, les membres ou les feuilles dans un plan corporel1. En raison de telles contraintes physiologiques et géométriques de la symétrie axiale, la plupart des embranchements de Bilateria- tels que les annélides, les arthropodes et les chordés- présentent une segmentation de leurs tissus embryonnaires (par exemple, ectoderme, mésoderme) antéro-postérieur.
Les embryons de vertébrés segmentent séquentiellement leur mésoderme paraxial le long de l’axe principal du corps en somites avec des intervalles, des comptes et des distributions de taille spécifiques à l’espèce. Malgré une telle robustesse parmi les embryons individuels au sein d’une espèce, la segmentation de la somite est polyvalente entre les espèces de vertébrés. La segmentation se produit dans un vaste régime d’intervalles de temps (de 25 min chez le poisson zèbre à 5 h chez l’homme), de tailles (de ~20 μm dans les somites de queue de poisson zèbre à ~200 μm dans les somites de tronc de souris) et de comptages (de 32 chez les poissons zèbres à ~300 chez les serpents de maïs)2. Plus intéressant encore, les embryons de poissons peuvent se développer dans une large gamme de températures (de ~ 20,5 ° C à 34 ° C pour le poisson zèbre) tout en gardant leurs somites intactes avec des distributions de taille appropriées en compensant à la fois les intervalles de segmentation et les vitesses d’allongement axiales. Au-delà de ces caractéristiques intéressantes, le poisson zèbre reste un organisme modèle utile pour étudier la segmentation chez les vertébrés en raison du développement externe, synchrone et transparent d’une plénitude d’embryons de frères et sœurs ainsi que de leurs outils génétiques accessibles. Du point de vue de la microscopie, les embryons de téléostéos se développent sur un jaune sphérique volumineux, étirant et arrondissant le tissu gazeux qui l’entoure (Figure 1A). En cet article, nous présentons une culture aplatie d’explant de tissu 3D pour des queues de poisson zèbre. Ce système d’explantation contourne les contraintes sphériques de la masse vitellin, permettant l’accès à l’imagerie vivante haute résolution d’embryons de poissons pour le modelage de somite.
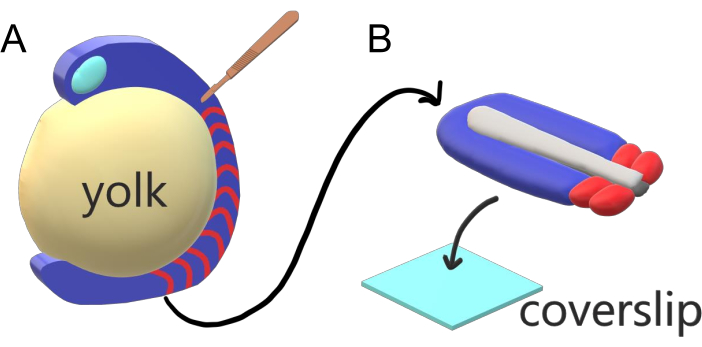
Figure 1: Système d’explantation à chambre coulissante pour les embryons de poisson zèbre. (A) Les embryons de poisson zèbre présentent des avantages pour l’imagerie vivante, tels que la transparence du tissu embryonnaire gastrulating (bleu), mais le tissu se forme autour d’une masse de jaune sphérique volumineuse (jaune) qui empêche l’imagerie proche de l’objectif et à haute résolution dans les embryons intacts. Les explants de queue peuvent être disséqués en commençant par un couteau microchirurgical (brun) coupé du tissu antérieur des somites (rouge) et en continuant à la frontière avec le jaune postérieurement. (B) Les explants de queue disséqués peuvent être placés sur une lamelle de couverture (bleu clair) dorsoventralement; garder le tissu neural (gris clair) sur le dessus et notochord (gris foncé) en bas. Veuillez cliquer ici pour voir une version plus grande de cette figure.
