Metameric segmentering av organismer används ofta i naturen. Upprepade strukturer är viktiga för funktionaliteten hos laterala organ som ryggkotor, muskler, nerver, kärl, lemmar eller löv i en kroppsplan1. Som ett resultat av sådana fysiologiska och geometriska begränsningar av axiell symmetri, de flesta phyla av Bilateria-såsom annelider, leddjur och ackord-utställning segmentering av deras embryonala vävnader (t.ex. ectoderm, mesoderm) antero-posteriorly.
Ryggradsdjursembryon segmenterar sekventiellt sin paraxiala mesoderm längs den stora kroppsaxeln till somiter med artspecifika intervall, antal och storleksfördelningar. Trots sådan robusthet bland enskilda embryon inom en art är somitsegmentering mångsidig mellan ryggradsdjur. Segmentering sker i en stor regim av tidsintervall (från 25 min i zebrafisk till 5 h hos människor), storlekar (från ~ 20 μm i svans somiter av zebrafisk till ~ 200 μm i stam somiter av möss) och räknas (från 32 i zebrafisk till ~ 300 i majsormar)2. Mer intressant är att fiskembryon kan utvecklas i ett brett spektrum av temperaturer (från ~ 20,5 ° C upp till 34 ° C för zebrafisk) samtidigt som de håller sina somiter intakta med rätt storleksfördelningar genom att kompensera för både segmenteringsintervall och axiella förlängningshastigheter. Utöver sådana intressanta egenskaper stannar zebrafisk som en användbar modellorganism för att studera segmentering i ryggradsdjur på grund av den externa, synkrona och transparenta utvecklingen av en plenitude av syskonembryon samt deras tillgängliga genetiska verktyg. Ur ett mikroskopiskt perspektiv utvecklas teleostembryon på en skrymmande sfärisk äggula, som sträcker och rundar den gastrulerande vävnaden runt den (figur 1A). I den här artikeln presenterar vi en tillplattad 3D-vävnad explant kultur för zebrafisk svansar. Detta explantsystem kringgår de sfäriska begränsningarna av äggula massa, vilket ger tillgång till högupplöst levande avbildning av fisk embryon för somite mönstring.
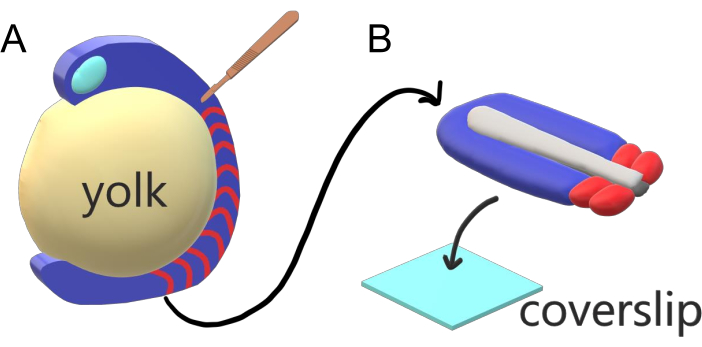
Figur 1:Slide Chamber Explant System for Zebrafish Embryos. (A) Zebrafiskembryon har fördelar för levande avbildning, såsom insyn i gastrulaterande embryonal vävnad (blå), men vävnaden bildas runt en skrymmande sfärisk äggulamassa (gul) som förhindrar nära objektiv, högupplöst avbildning i intakta embryon. Svans explanter kan dissekeras börjar med en mikrokirurgisk kniv (brun) skuren från vävnaden främre av somiter (röd) och fortsätter vid gränsen till äggulan bakre. B)Dissekerade svansexplanter kan placeras på ett täckglas (ljusblått) dorsoventrally. hålla neural vävnad (ljusgrå) på toppen och notochord (mörkgrå) längst ner. Klicka här om du vill visa en större version av den här figuren.
