Som en live-celle DFS teknik, BFP ingeniører en menneskelig rød blodlegemer (RBC; Figur 1) til en ultrafølsom og tunable krafttransducer med et kompatibelt fjederkonstantområde ved 0,1-3 pN/nm1,2,3. For at sondere ligand-receptor interaktion, BFP muliggør DFS målinger på ~ 1 pN (10-12 N), ~ 3 nm (10-9 m), og ~ 0,5 ms (10-3 s) i kraft, rumlig og tidsmæssig opløsning4,5. Dens eksperimentelle konfiguration består af to modsatrettede mikropipetter, nemlig sonden og målet. Probe mikropipetten aspireerer en RBC og en perle er limet på sit højdepunkt via en biotin-streptavidin interaktion. Perlen er belagt med ligand af interesse (figur 1A). Target-mikropipetten aspireerer enten en celle eller en perle, der bærer receptoren af interesse, svarende til perlecelle –figur 1B) og perleperle (Figur 1C) tilstande, henholdsvis5.
BFP konstruktion, samling og DFS eksperimentelle protokoller blev beskrevet i detaljer tidligere1,6. Kort fortalt består en BFP-berøringscyklus af 5 faser: Tilgang, Hindring, Kontakt, Tilbagetrækning og Dissociate (Figur 1D). Den vandrette RBC-spidsposition er angivet som ΔxRBC. I begyndelsen er den ustrøede (nul-kraft) RBC deformation ΔxRBC 0 (Tabel 1). Målet køres derefter af en piezotranslator for at gribe ind og trække sig tilbage fra sondeperen (Figur 1D). RBC-sonden komprimeres først af Target med negativ RBC-deformation ΔxRBC < 0. I en Bond-hændelse skifter tilbagetrækningsstadiet fra et kompressivt til en trækfase med positiv RBC-deformation ΔxRBC > 0 (Figur 2C og D). Ifølge Hookes lov kan BFP’s bærekraft måles som F = kRBC × ΔxRBC, hvor kRBC ( tabel1) er BFP’s RBC-fjederkonstant. Ved lim brud og afslutningen af en touch cyklus, sonden perle vender tilbage til nul-kraft position med ΔxRBC = 0 (Figur 1D).
For at bestemme kRBCmåler og registrerer vi radierne i sondemikropipettens indre åbning (Rp), RBC (R0) og det cirkulære kontaktområde (Rc) mellem RBC og sondeperen ( Figur1A). Derefter beregnes kRBC i henhold til Evans model (Eq. 1)7,8 ved hjælp af et LabVIEW-program, der fungerer som et virtuelt instrument (VI) til at betjene BFP ( FigurS1A)8,9.
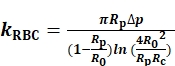 (Eq. 1)
(Eq. 1)
Med en BFP etableret og DFS rå data opnået, hermed præsenterer vi, hvordan man analyserer foråret konstant af ligand-receptor par eller celler. DFS-rådata om samspillet mellem det glycosylerede protein Thy-1 og K562-cellen med integrin α5β1 (Thy-1-α5β1; Figur 3A og 3B)10 og fibrinogen og perlebelagt integrin αIIbβ3 (FGN-αIIbβ3; Figur 3C) 11,12 er blevet brugt til at demonstrere henholdsvis perlecelle- og perleperleanalysetilstandene.
BFP eksperimentel forberedelse
Yderligere oplysninger om BFP’s eksperimentelle forberedelse og instrumentering henvises til de tidligere offentliggjorte protokoller3. Kort sagt er human RBC blevet biotinyleret ved hjælp af Biotin-PEG3500-NHS i kulstof /bicarbonatbufferen. Proteiner af interesse er blevet kovalent koblet til borosilicate glasperler ved hjælp af MAL-PEG3500-NHS i fosfat buffer. For at fastgøres til den biotinylerede RBC er sondeperen også belagt med streptavidin (SA) ved hjælp af MAL-SA. Se tabellen over materialer og tabel 2.
For at samle BFP(figur 1, til venstre)vil den tredje mikropipette , der kaldes ‘Hjælper’, blive brugt til at levere sondeperen og lime den fast til RBC’s spids1,3. Den kovalente interaktion mellem sa-coated sondeper og biotinyleret RBC er meget stærkere end ligandreceptorbindingen af interesse. Således kan Dissociate-stadiet fortolkes som ligand-receptorbindingsbrud snarere end løsrivelse af Probe perle fra RBC.
