Als een live-cell DFS-techniek ontwikkelt BFP een menselijke rode bloedcel (RBC; Figuur 1) in een ultragevoelige en afstembare krachtopnemer met een compatibel veerconstantebereik bij 0,1-3 pN/nm1,2,3. Om ligand-receptorinteractie te onderzoeken, maakt BFP DFS-metingen mogelijk bij ~ 1 pN (10-12 N), ~ 3 nm(10 -9 m) en ~ 0,5 ms (10-3 s) in kracht, ruimtelijke en temporele resolutie4,5. De experimentele configuratie bestaat uit twee tegengestelde micropipettes, namelijk de Probe en het doelwit. De Probe micropipette aspirateert een RBC en een kraal wordt aan de top gelijmd via een biotine-streptavidin interactie. De kraal is bedekt met het ligand van belang (figuur 1A). De doelmicropipette aspirateert een cel of een kraal met de receptor van belang, overeenkomend met de modi Bead-Cell (Figuur 1B) en Bead-Bead (Figuur 1C) respectievelijk5.
BFP constructie, assemblage en de DFS experimentele protocollen werden eerder in detail beschreven1,6. In het kort bestaat een BFP-aanraakcyclus uit 5 fasen: Approach, Impinge, Contact, Retract en Dissociate(Figuur 1D). De horizontale RBC-toppositie wordt aangeduid als ΔxRBC. In het begin is de onbelaste (nulkracht) RBC-vervorming ΔxRBC 0 (tabel 1). Het doel wordt vervolgens aangedreven door een piëzotranslator om de sondekraal te impeneren en terug te trekken(figuur 1D). De RBC-sonde wordt eerst gecomprimeerd door het doel met negatieve RBC-vervorming ΔxRBC < 0. In een Bond-gebeurtenis gaat de Retract-fase over van een druk- naar een trekfase met positieve RBC-vervorming ΔxRBC > 0 (figuur 2C en D). Volgens de wet van Hooke kan de BFP-draagkracht worden gemeten als F = kRBC × ΔxRBC, waarbij kRBC ( tabel1) de RBC-veerconstante van de BFP is. Na het scheuren van de binding en het voltooien van de one-touch cyclus, keert de sondekraal terug naar de nulkrachtpositie met ΔxRBC = 0 (Figuur 1D).
Om de kRBCte bepalen, meten en registreren we de radii van de binnenste opening van de sondemicropipette (Rp), de RBC (R0) en het cirkelvormige contactgebied (Rc) tussen de RBC en de sondekraal ( Figuur1A). Vervolgens wordt kRBC berekend volgens het model van Evan (Eq. 1)7,8 met behulp van een LabVIEW-programma dat fungeert als een virtueel instrument (VI) om de BFP te bedienen (Figuur S1A)8,9.
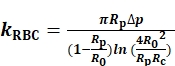 (Eq. 1)
(Eq. 1)
Met een BFP vastgesteld en DFS ruwe gegevens verkregen, presenteren we hierbij hoe de veerconstante van ligand-receptorpaar of cellen te analyseren. De DFS ruwe gegevens over de interactie van het geglycosyleerde eiwit Thy-1 en K562 celdragende integrine α5β1 (Thy-1-α5β1; Figuren 3A en 3B)10 en die van fibrinogeen en met kralen bekleed integrine αIIbβ3 (FGN-αIIbβ3; Figuur 3C) 11,12 zijn gebruikt om respectievelijk de Bead-Cell en Bead-Bead analysemodi te demonstreren.
BFP Experimentele Voorbereiding
Voor meer informatie over de experimentele voorbereiding en instrumentatie van BFP verwijzen wij u naar de eerder gepubliceerde protocollen3. Kortom, humane RBC is gebiotinyleerd met behulp van de Biotine-PEG3500-NHS in de koolstof / bicarbonaatbuffer. Interessante eiwitten zijn covalent gekoppeld aan de borosilicaatglasparels met behulp van MAL-PEG3500-NHS in de fosfaatbuffer. Om zich aan de gebiotinyleerde RBC te hechten, wordt de sondekraal ook bedekt met streptavidin (SA) met behulp van de MAL-SA. Zie de tabel met materialen en tabel 2.
Om de BFP te assembleren(figuur 1, links),zal de derde micropipette met de term ‘Helper’ worden gebruikt om de sondekraal af te leveren en te lijmen op de top1,3vande RBC. De covalente interactie tussen de SA-gecoate sondekraal en gebiotinyleerde RBC is veel sterker dan de ligand-receptorbinding van belang. Het dissociate stadium kan dus worden geïnterpreteerd als de ligand-receptorbindingsruptuur in plaats van de loslating van probekraal van de RBC.
