En tant que technique DFS à cellules vivantes, BFP conçoit un globule rouge humain (RBC; Figure 1) dans un transducteur de force ultrasensible et accordable avec une plage constante de ressort compatible à 0,1-3 pN/nm1,2,3. Pour sonder l’interaction ligand-récepteur, BFP permet des mesures DFS à ~1 pN(10 -12 N), ~3 nm(10 -9 m) et ~0,5 ms (10-3 s) en force, résolution spatiale et temporelle4,5. Sa configuration expérimentale se compose de deux micropipettes opposées, à savoir la sonde et la cible. La micropipette Probe aspire un globules biologiques et une perle est collée à son sommet via une interaction biotine-streptavidine. La perle est recouverte du ligand d’intérêt (Figure 1A). La micropipette cible aspire soit une cellule, soit une perle portant le récepteur d’intérêt, correspondant aux modes Bead-Cell(Figure 1B)et Bead-Bead(Figure 1C),respectivement5.
La construction BFP, l’assemblage et les protocoles expérimentaux DFS ont été décrits en détail précédemment1,6. En bref, un cycle tactile BFP se compose de 5 étapes : Approcher, Impinge, Contact, Retract et Dissocier(Figure 1D). La position horizontale de l’apex RBC est notée ΔxRBC. Au début, la déformation RBC non étayée (force nulle) ΔxRBC est 0(tableau 1). La cible est ensuite entraînée par un piézotranslateur pour empiéter et se rétracter sur le cordon de la sonde(Figure 1D). La sonde RBC est d’abord comprimée par la cible avec une déformation RBC négative ΔxRBC < 0. Dans un cas de liaison, l’étage de rétraction passe d’une phase de compression à une phase de traction avec une déformation RBC positive ΔxRBC > 0(Figure 2C et D). Selon la loi de Hooke, la force de roulement BFP peut être mesurée comme F = kRBC × ΔxRBC, où kRBC ( Tableau1) est la constante de ressort RBC du BFP. À la rupture de la liaison et à la fin d’un cycle tactile, le bille de sonde revient à la position de force nulle avec ΔxRBC = 0(Figure 1D).
Pour déterminer le kRBC,nous mesurons et enregistrons les rayons de l’orifice interne de la micropipette de sonde (Rp), du RBC (R0) et de la zone de contact circulaire (Rc) entre le RBC et le bille de sonde ( Figure1A). Ensuite, kRBC est calculé selon le modèle d’Evan (Eq. 1)7,8 à l’aide d’un programme LabVIEW qui agit comme un instrument virtuel (VI) pour faire fonctionner le BFP(Figure S1A)8,9.
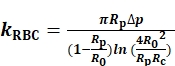 (Eq. 1)
(Eq. 1)
Avec un BFP établi et des données brutes DFS obtenues, nous présentons ici comment analyser la constante de ressort de la paire ligand-récepteur ou des cellules. Les données brutes DFS sur l’interaction de la protéine glycosylée Thy-1 et de la cellule K562 portant l’intégrine α5β1 (Thy-1-α5β1; Figures 3A et 3B)10 et celle de l’intégrine enduite de fibrinogène et de billes αIIbβ3 (FGN-αIIbβ3; Figure 3C) 11,12 ont été utilisés pour démontrer les modes d’analyse Bead-Cell et Bead-Bead, respectivement.
Préparation expérimentale BFP
Pour plus de détails sur la préparation expérimentale et l’instrumentation du BFP, veuillez vous référer aux protocoles précédemment publiés3. En bref, les globules biologiques humains ont été biotinylés à l’aide du Biotin-PEG3500-NHS dans le tampon carbone/bicarbonate. Les protéines d’intérêt ont été couplées de manière covalente aux billes de verre borosilicate en utilisant MAL-PEG3500-NHS dans le tampon de phosphate. Pour se fixer au globule rouge biotinylé, la bille de sonde est également recouverte de streptavidine (SA) à l’aide du MAL-SA. Veuillez consulter le tableau des matériaux et le tableau 2.
Pour assembler le BFP(Figure 1, à gauche),la troisième micropipette appelée « Helper » sera utilisée pour délivrer la bille de sonde et la coller à l’apex du RBC1,3. L’interaction covalente entre la bille de sonde revêtue de SA et les globules rouges biotinylés est beaucoup plus forte que la liaison ligand-récepteur d’intérêt. Ainsi, l’étape dissociée peut être interprétée comme la rupture de la liaison ligand-récepteur plutôt que le détachement de la perle de sonde du globule rouge.
