La voie endocytaire non spécifique de la macropinocytose permet aux cellules d’internaliser une variété de composants extracellulaires, y compris les nutriments, les protéines, les antigènes et les agents pathogènes, grâce à l’absorption en vrac du liquide extracellulaire et de ses constituants1. Bien qu’importante pour la biologie de nombreux types de cellules, la voie de la macropinocytose est décrite comme jouant un rôle essentiel dans la biologie tumorale, où, grâce à l’absorption macropinocytaire, les cellules tumorales sont capables de survivre et de proliférer en présence d’un microenvironnement appauvri en nutriments2,3. L’absorption des macromolécules extracellulaires, y compris l’albumine et la matrice extracellulaire, et des débris cellulaires nécrotiques, fournit une source alternative de nutriments pour la production de biomasse en créant des acides aminés, des sucres, des lipides et des nucléotides par le biais du catabolisme de cargaison médié par la fusion de macropinosomes et de lysosomes4,5,6,7,8.
L’induction et la régulation de la macropinocytose sont complexes et peuvent varier en fonction du contexte cellulaire. Jusqu’à présent, plusieurs inducteurs de la macropinocytose ont été identifiés et comprennent des ligands, tels que le facteur de croissance épidermique (EGF), le facteur de croissance dérivé des plaquettes (PDGF), la galectine-3 et Wnt3A9,10,11,12,13. En outre, les conditions de culture qui imitent le microenvironnement tumoral peuvent déclencher l’activation de la voie. Les tumeurs de l’adénocarcinome canalaire pancréatique (PDAC) sont privées de nutriments, en particulier pour l’acide aminé glutamine, ce qui fait que les cellules cancéreuses et les fibroblastes associés au cancer (CAF) dépendent de la macropinocytose pour la survie7,13,14,15. De plus, les stress tumoraux, tels que l’hypoxie et le stress oxydatif, peuvent activer cette voie de piégeage16. En plus des nombreux influenceurs extrinsèques qui peuvent induire la macropinocytose, une variété de voies intracellulaires contrôlent la formation de macropinosomes. La transformation oncogène médiée par Ras est suffisante pour initier la machinerie macropinocytaire, et plusieurs types de cancer présentent une macropinocytose constitutive oncogène pilotée par Ras4,5,9,17. Alternativement, l’activation ras de type sauvage et les voies indépendantes de Ras ont été identifiées pour activer la macropinocytose dans les cellules cancéreuses et les CAF10,11,15,18. L’utilisation de divers modèles in vitro en combinaison avec des traitements inhibiteurs a abouti à l’identification de plusieurs modulateurs de macropinocytose, qui comprennent des échangeurs sodium-hydrogène, la petite GTPase Rac1, la phosphoinositide 3-kinase (PI3K), la kinase activée par p21 (Pak) et la protéine kinase activée par AMP (AMPK)4,13,15 . Cependant, étant donné la multitude de facteurs et de conditions décrits qui régulent la macropinocytose, il est concevable que beaucoup plus de modulateurs et de stimuli restent inconnus. L’identification de nouveaux modulateurs et stimuli peut être facilitée par l’évaluation automatisée d’une multitude de conditions dans une seule expérience. Cette méthodologie peut faire la lumière sur les facteurs impliqués dans la formation de macropoinosomes et peut permettre la découverte de nouvelles petites molécules ou de produits biologiques qui ciblent cette voie.
Ici, nous avons adapté notre protocole précédemment établi pour déterminer l’étendue de la macropinocytose dans les cellules cancéreuses in vitro à un format de microplaque à 96 puits et à une imagerie et une quantification automatisées19,20. Ce protocole est basé sur la microscopie fluorescente, qui est devenue une norme dans le domaine pour déterminer la macropinocytose in vitro et in vivo4,5,6,7,9,10,11,12,13,15,16,17,18, 19,20,21,22. Les macropoinosomes peuvent être distingués des autres voies endocytaires par leur capacité à internaliser de grandes macromolécules, telles que le dextran de haut poids moléculaire (c.-à-d. 70 kDa)2,3,4,20,21,22,23. Ainsi, les macropinosomes peuvent être définis par l’absorption de fluorophores fluorophores marqués extracellulairement 70 kDa dextran. En conséquence, les vésicules macropinocytaires se manifestent sous forme d’amas intracellulaires de puncta fluorescents de tailles allant de 0,2 à 5 μm. Ces puncta peuvent être imagés au microscope et ensuite quantifiés pour déterminer l’étendue de la macropinocytose dans la cellule – « l’indice macropinocytaire ».
Dans ce protocole, les étapes essentielles pour visualiser les macropinosomes dans les cellules adhérentes in vitro sur une microplaque de 96 puits et des couvercles à l’aide d’équipements de laboratoire standard sont décrites (Figure 1). En outre, les instructions pour automatiser l’acquisition d’images et la quantification de l’indice macropinocytaire à l’aide d’un lecteur de plaques multimode d’imagerie cellulaire sont fournies. Cette automatisation réduit le temps, les coûts et les efforts par rapport à nos protocoles décrits précédemment19,20. En outre, il évite l’acquisition et l’analyse d’images involontairement biaisées et améliore ainsi la reproductibilité et la fiabilité. Cette méthode peut facilement être adaptée à différents types de cellules ou de lecteurs de plaques ou être utilisée pour déterminer d’autres caractéristiques des macropoinosomes, telles que la taille, le nombre et l’emplacement. La méthode décrite ici est particulièrement adaptée au dépistage des conditions de culture cellulaire qui induisent la macropinocytose, à l’identification de nouveaux modulateurs ou à l’optimisation des concentrations médicamenteuses d’inhibiteurs connus.
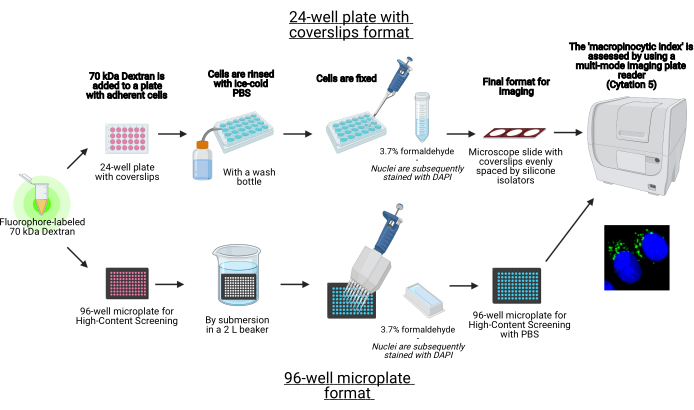
Figure 1 : Schéma du test automatisé pour déterminer l’indice macropinocytaire dans les cellules adhérentes. Créé à l’aide de BioRender. Veuillez cliquer ici pour voir une version agrandie de cette figure.
