Den ikke-spesifikke endokytiske banen til makropinocytose gjør det mulig for celler å internalisere en rekke ekstracellulære komponenter, inkludert næringsstoffer, proteiner, antigener og patogener, gjennom bulkopptak av ekstracellulær væske og dens bestanddeler1. Selv om det er viktig for biologien til mange celletyper, er makropinocytosebanen i økende grad beskrevet for å spille en viktig rolle i tumorbiologien, hvor tumorceller gjennom makropinocytisk opptak er i stand til å overleve og spre seg i nærvær av et næringsfattig mikromiljø2,3. Opptaket av ekstracellulære makromolekyler, inkludert albumin og ekstracellulær matrise, og nekrotisk celleavfall, gir en alternativ næringskilde for biomasseproduksjon ved å lage aminosyrer, sukker, lipider og nukleotider gjennom makropinosom og lysosomet fusjonsmediert lastkaboisme4,5,6,7,8.
Induksjonen og reguleringen av makropinocytose er kompleks og kan variere avhengig av cellulær kontekst. Så langt har flere indusere av makropinocytose blitt identifisert og inkluderer ligander, for eksempel epidermal vekstfaktor (EGF), blodplate-avledet vekstfaktor (PDGF), galectin-3 og Wnt3A9,10,11,12,13. I tillegg kan dyrkingsforhold som etterligner tumormikromiljøet utløse aktivering av banen. Bukspyttkjertelkanal adenokarsinom (PDAC) svulster er næringsfattige, spesielt for aminosyre glutamin, noe som fører til at både kreftceller og kreftrelaterte fibroblaster (CAFs) er avhengige av makropinocytose for overlevelse7,13,14,15. Videre kan tumorspenninger, som hypoksi og oksidativt stress, aktivere denne scavenging pathway16. I tillegg til de mange ekstrinsiske påvirkerne som kan indusere makropinocytose, kontrollerer en rekke intracellulære veier makropinosomdannelse. Onkogen Ras-mediert transformasjon er tilstrekkelig til å initiere det makropinocytiske maskineriet, og flere krefttyper utviser onkogen Ras-drevet konstituerende makropinocytose4,5,9,17. Alternativt er villtype Ras-aktivering og Ras-uavhengige veier identifisert for å aktivere makropinocytose i kreftceller og CAFs10,11,15,18. Bruken av ulike in vitro-modeller i kombinasjon med inhibitorbehandlinger har resultert i identifisering av flere makropinocytosemodulatorer, som inkluderer natrium-hydrogenvekslere, den lille GTPase Rac1, fosfoinositid 3-kinase (PI3K), p21-aktivert kinase (Pak) og AMP-aktivert proteinkinase (AMPK) 4,13,15 . Men gitt mangfoldet av beskrevne faktorer og forhold som regulerer makropinocytose, er det tenkelig at mange flere modulatorer og stimuli forblir uoppdaget. Identifisering av nye modulatorer og stimuli kan lettes ved automatisert vurdering av en rekke forhold i et enkelt eksperiment. Denne metoden kan belyse faktorene som er involvert i makropinosomdannelse og kan tillate oppdagelsen av nye små molekyler eller biologer som retter seg mot denne veien.
Her har vi tilpasset vår tidligere etablerte protokoll for å bestemme omfanget av makropinocytose i kreftceller in vitro til et 96-brønns mikroplateformat og automatisert avbildning og kvantifisering19,20. Denne protokollen er basert på fluorescerende mikroskopi, som har blitt en standard i feltet for å bestemme makropinocytose in vitro og in vivo4,5,6,7,9,10,11,12,13,15,16,17,18, 19,20,21,22. Makropinosomer kan skille seg fra andre endokytiske veier gjennom deres evne til å internalisere store makromolekyler, som høymolekylær dextran (dvs. 70 kDa)2,3,4,20,21,22,23. Dermed kan makropinosomer defineres gjennom opptak av ekstracellulært administrert fluorofor-merket 70 kDa dextran. Som et resultat manifesterer makropinocytiske vesikler som intracellulære klynger av fluorescerende puncta med størrelser fra 0,2-5 μm. Disse puncta kan mikroskopisk avbildes og deretter kvantifiseres for å bestemme omfanget av makropinocytose i cellen – ‘den makropinocytiske indeksen’.
I denne protokollen er de viktigste trinnene for å visualisere makropinosomer i adherentceller in vitro på en 96-brønns mikroplate og deksler ved hjelp av standard laboratorieutstyr beskrevet (figur 1). I tillegg er instruksjonene for å automatisere bildeanskaffelsen og kvantifiseringen av den makropinocytiske indeksen ved hjelp av en celleavbildningsplateleser i flere moduser gitt. Denne automatiseringen reduserer tid, kostnader og krefter sammenlignet med våre tidligere beskrevne protokoller19,20. I tillegg unngår den utilsiktet partisk bildeinnsamling og analyse og forbedrer dermed reproduserbarhet og pålitelighet. Denne metoden kan enkelt tilpasses ulike celletyper eller platelesere eller brukes til å bestemme alternative makropinosomfunksjoner, for eksempel størrelse, antall og plassering. Den heri beskrevne metoden er spesielt egnet for screening av cellekulturforhold som induserer makropinocytose, identifisering av nye modulatorer eller optimalisering av legemiddelkonsentrasjoner av kjente inhibitorer.
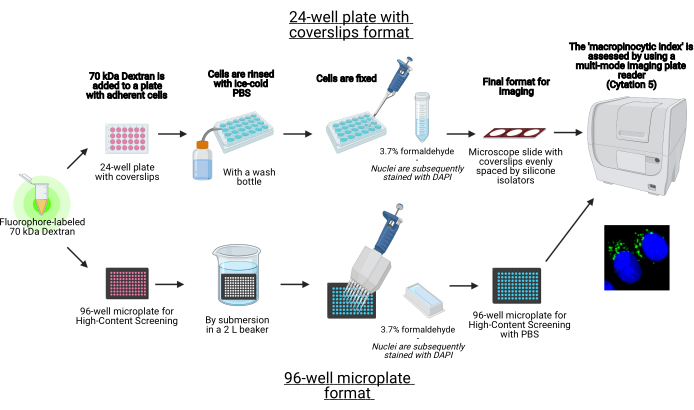
Figur 1: Skjematisk for den automatiserte analysen for å bestemme den ‘makropinocytiske indeksen’ i tilhengerceller. Opprettet ved hjelp av BioRender. Klikk her for å se en større versjon av denne figuren.
