منذ فترة طويلة تم التغاضي عن Strongyloides stercoralis كممرض بشري مهم مقارنة بالديدان الخطافية المعترف بها على نطاق واسع والدودة المستديرة Ascaris lumbricoides1. غالبا ما قللت الدراسات السابقة لعبء الدودة بشدة من انتشار S. stercoralis بسبب الحساسية المنخفضة لطرق التشخيص الشائعة ل S. stercoralis2. في السنوات الأخيرة ، قدرت الدراسات الوبائية القائمة على أدوات التشخيص المحسنة أن الانتشار الحقيقي لعدوى S. stercoralis أعلى بكثير مما تم الإبلاغ عنه سابقا ، أي ما يقرب من 610 ملايين شخص في جميع أنحاء العالم2.
كل من S. stercoralis وأنواع Strongyloides الأخرى ، بما في ذلك طفيلي الفئران المرتبط ارتباطا وثيقا ونموذج المختبر المشترك S. ratti ، لديهم دورة حياة غير عادية مفيدة للدراسات الجينومية التجريبية لأنها تتكون من أجيال طفيلية وحرة (بيئية)3 (الشكل 1). على وجه التحديد ، يمكن لكل من S. stercoralis و S. ratti التنقل عبر جيل واحد من العيش الحر. يتكون جيل العيش الحر من يرقات ما بعد الطفيلية التي تتطور إلى ذكور وإناث بالغين يعيشون بحرية. تتطور جميع ذرية البالغين الذين يعيشون بحرية إلى يرقات معدية ، والتي يجب أن تصيب المضيف لمواصلة دورة الحياة. علاوة على ذلك ، يمكن التلاعب بهذا الجيل البيئي أو الحي الحر تجريبيا في المختبر. نظرا لأن البالغين الذين يعيشون بحرية في Strongyloides و C. elegans البالغين يشتركون في مورفولوجيا مماثلة ، يمكن تكييف تقنيات مثل الحقن المجهري داخل الغدد التناسلية التي تم تطويرها في الأصل ل C. elegans للاستخدام مع Strongyloides 4,5 البالغين الذين يعيشون بحرية. في حين يتم إدخال الحمض النووي بشكل عام في الإناث البالغات اللواتي يعشن بحرية ، يمكن حقن كل من الذكور والإناث من Strongyloides 6. وبالتالي ، تتوفر أدوات الجينوم الوظيفية لاستجواب العديد من جوانب بيولوجيا Strongyloides. تفتقر الديدان الخيطية الطفيلية الأخرى إلى جيل يعيش بحرية ، ونتيجة لذلك ، فهي ليست قابلة بسهولة للتقنيات الجينومية الوظيفية3.
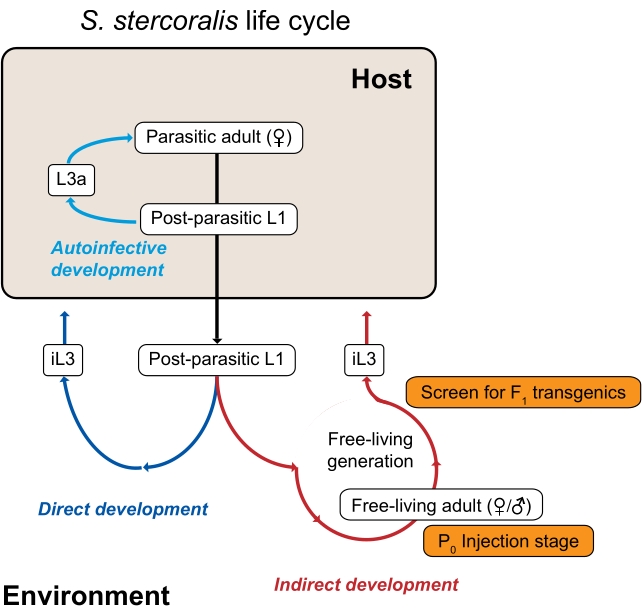
الشكل 1: دورة حياة سترونغيلويدات ستيركوراليس. تعيش الإناث الطفيلية S. stercoralis في الأمعاء الدقيقة لمضيفيها من الثدييات (البشر ، الرئيسيات غير البشرية ، الكلاب). تتكاثر الإناث الطفيلية عن طريق التوالد العذري وتضع البيض داخل الأمعاء الدقيقة. يفقس البيض بينما لا يزال داخل المضيف إلى يرقات ما بعد الطفيلية ، والتي يتم تمريرها بعد ذلك إلى البيئة مع البراز. إذا كانت يرقات ما بعد الطفيليات من الذكور ، فإنها تتطور إلى ذكور بالغين يعيشون بحرية. إذا كانت يرقات ما بعد الطفيلية أنثى ، فيمكنها إما أن تتطور إلى إناث بالغات يعشن بحرية (تطور غير مباشر) أو يرقات معدية في المرحلة الثالثة (iL3s ؛ تطور مباشر). الذكور والإناث الذين يعيشون بحرية يتكاثرون جنسيا لخلق ذرية مقيدة لتصبح iL3s. في ظل ظروف معينة ، يمكن أن يخضع S. stercoralis أيضا للعدوى الذاتية ، حيث تبقى بعض يرقات ما بعد الطفيلية داخل الأمعاء المضيفة بدلا من المرور إلى البيئة في البراز. يمكن أن تتطور هذه اليرقات إلى يرقات ذاتية العدوى (L3a) داخل المضيف ، وتخترق جدار الأمعاء ، وتهاجر عبر الجسم ، وتعود في النهاية إلى الأمعاء لتصبح بالغين تناسليين. دورة حياة S. ratti متشابهة ، باستثناء أن S. ratti يصيب الفئران وليس لديه دورة عدوى ذاتية. الجيل البيئي هو المفتاح لاستخدام أنواع Strongyloides للدراسات الوراثية. يمكن حقن الإناث البالغات اللواتي يعشن بحرية (P0) بشكل مجهري ؛ ذريتهم ، والتي ستصبح جميعها iL3s ، هي المعدلة وراثيا F1 المحتملة. وقد عدل هذا الرقم من كاستيليتو وآخرين. 3. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
تشترك S. stercoralis في العديد من جوانب بيولوجيتها مع الديدان الخيطية الطفيلية البشرية المعدية المعوية الأخرى ، بما في ذلك غزو المضيف وتعديل المناعة المضيفة. على سبيل المثال ، تصيب الديدان الخطافية البشرية الطفيلية في الأجناس Necator و Ancylostoma أيضا عن طريق اختراق الجلد ، وتتنقل بالمثل عبر الجسم ، وتقيم في النهاية كبالغين طفيليين في الأمعاء الدقيقة7. وبالتالي ، من المحتمل أن تستخدم العديد من الديدان الخيطية المعدية المعوية السلوكيات الحسية الشائعة وتقنيات التهرب المناعي. ونتيجة لذلك ، فإن المعرفة المستقاة من Strongyloides ستكمل النتائج في الديدان الخيطية الأخرى الأقل قابلية للسحب وراثيا وتؤدي إلى فهم أكثر اكتمالا لهذه الطفيليات المعقدة والمهمة.
يحدد بروتوكول الحقن المجهري هذا طريقة إدخال الحمض النووي في الإناث البالغات اللواتي يعشن بحرية في Strongyloides لصنع ذرية معدلة وراثيا ومتحولة. يتم وصف متطلبات صيانة السلالة ، بما في ذلك التوقيت التنموي للديدان البالغة للحقن المجهري وجمع النسل المعدل وراثيا. يتم تضمين البروتوكولات وعرض توضيحي لتقنية الحقن المجهري الكاملة ، إلى جانب بروتوكولات زراعة وفحص النسل المعدل وراثيا ، إلى جانب قائمة بجميع المعدات والمواد الاستهلاكية اللازمة.
