Strongyloides stercoralis è stato a lungo trascurato come un importante agente patogeno umano rispetto agli anchilostomi più ampiamente riconosciuti e al nematode Ascaris lumbricoides1. Precedenti studi sul carico di vermi spesso hanno gravemente sottovalutato la prevalenza di S. stercoralis a causa della bassa sensibilità dei metodi diagnostici comuni per S. stercoralis2. Negli ultimi anni, studi epidemiologici basati su strumenti diagnostici migliorati hanno stimato che la vera prevalenza delle infezioni da S. stercoralis è molto più alta di quanto precedentemente riportato, circa 610 milioni di persone in tutto il mondo2.
Sia S. stercoralis che altre specie di Strongyloides, tra cui il parassita di ratto strettamente correlato e il comune modello di laboratorio S. ratti, hanno un ciclo di vita insolito che è vantaggioso per gli studi genomici sperimentali perché consiste di generazioni sia parassitarie che libere viventi (ambientali)3 (Figura 1). In particolare, sia S. stercoralis che S. ratti possono attraversare un’unica generazione di vita libera. La generazione a vita libera è costituita da larve post-parassitarie che si sviluppano in maschi e femmine adulti che vivono liberi; tutta la progenie degli adulti che vivono liberi si sviluppa in larve infettive, che devono infettare un ospite per continuare il ciclo di vita. Inoltre, questa generazione ambientale o di vita libera può essere manipolata sperimentalmente in laboratorio. Poiché gli adulti Strongyloides a vita libera e gli adulti C. elegans condividono una morfologia simile, tecniche come la microiniezione intragonennale originariamente sviluppate per C. elegans possono essere adattate per l’uso con Strongyloides 4,5 adulti a vita libera. Mentre il DNA viene generalmente introdotto nelle femmine adulte che vivono liberamente, sia i maschi che le femmine di Strongyloides possono essere microiniettati6. Pertanto, sono disponibili strumenti genomici funzionali per interrogare molti aspetti della biologia di Strongyloides. Altri nematodi parassiti mancano di una generazione di vita libera e, di conseguenza, non sono così facilmente suscettibili alle tecniche genomiche funzionali3.
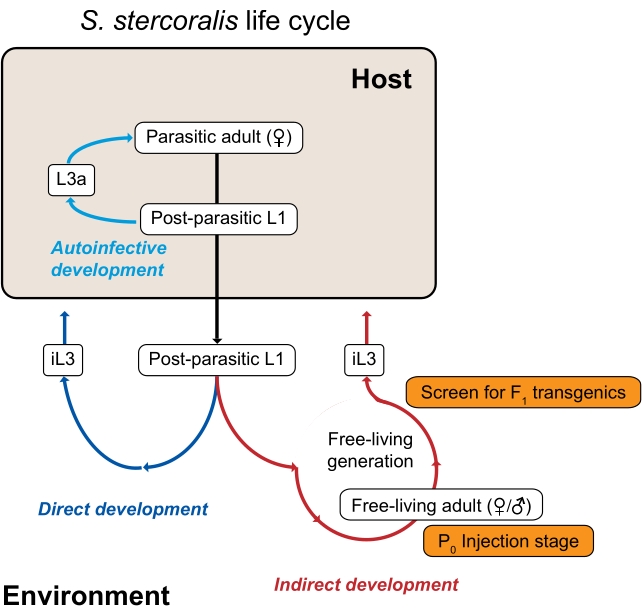
Figura 1: Il ciclo di vita di Strongyloides stercoralis. Le femmine parassitarie di S. stercoralis abitano l’intestino tenue dei loro ospiti mammiferi (umani, primati non umani, cani). Le femmine parassitarie si riproducono per partenogenesi e depongono le uova all’interno dell’intestino tenue. Le uova si schiudono mentre sono ancora all’interno dell’ospite in larve post-parassitarie, che vengono poi passate nell’ambiente con le feci. Se le larve post-parassitarie sono maschi, si sviluppano in maschi adulti che vivono liberamente. Se le larve post-parassitarie sono femmine, possono svilupparsi in femmine adulte a vita libera (sviluppo indiretto) o larve infettive di terzo stadio (iL3s; sviluppo diretto). I maschi e le femmine che vivono liberi si riproducono sessualmente per creare progenie che sono costretti a diventare iL3. In determinate condizioni, S. stercoralis può anche subire l’autoinfezione, in cui alcune delle larve post-parassitarie rimangono all’interno dell’intestino ospite piuttosto che passare nell’ambiente nelle feci. Queste larve possono svilupparsi in larve autoinfettive (L3a) all’interno dell’ospite, penetrare attraverso la parete intestinale, migrare attraverso il corpo e alla fine tornare all’intestino per diventare adulti riproduttivi. Il ciclo di vita di S. ratti è simile, tranne che S. ratti infetta i ratti e non ha un ciclo autoinfettivo. La generazione ambientale è la chiave per l’utilizzo delle specie di Strongyloides per studi genetici. Le femmine adulte a vita libera (P0) possono essere microintetate; la loro progenie, che diventerà tutta iL3, sono i potenziali transgenici F1. Questa figura è stata modificata da Castelletto et al. 3. Fare clic qui per visualizzare una versione più grande di questa figura.
S. stercoralis condivide molti aspetti della sua biologia con altri nematodi gastrointestinali parassiti umani, tra cui l’invasione dell’ospite e la modulazione immunitaria dell’ospite. Ad esempio, gli anchilostomi umani-parassiti nei generi Necator e Ancylostoma infettano anche la penetrazione della pelle, navigano in modo simile attraverso il corpo e, infine, risiedono come adulti parassiti nell’intestino tenue7. Pertanto, molti nematodi gastrointestinali probabilmente utilizzano comportamenti sensoriali comuni e tecniche di evasione immunitaria. Di conseguenza, le conoscenze raccolte da Strongyloides completeranno i risultati in altri nematodi meno trattabili geneticamente e porteranno a una comprensione più completa di questi parassiti complessi e importanti.
Questo protocollo di microiniezione delinea il metodo per introdurre il DNA nelle femmine adulte viventi libere di Strongyloides per produrre progenie transgenica e mutante. Vengono descritti i requisiti di mantenimento del ceppo, compresi i tempi di sviluppo dei vermi adulti per le microiniezioni e la raccolta della progenie transgenica. Sono inclusi protocolli e una dimostrazione della tecnica completa di microiniezione, insieme ai protocolli per la coltura e lo screening della progenie transgenica, insieme a un elenco di tutte le attrezzature e i materiali di consumo necessari.
