Strongyloides stercoralis a longtemps été négligé comme un agent pathogène humain important par rapport aux ankylostomes plus largement reconnus et au ver rond Ascaris lumbricoides1. Des études antérieures sur la charge de vers ont souvent gravement sous-estimé la prévalence de S. stercoralis en raison de la faible sensibilité des méthodes de diagnostic courantes pour S. stercoralis2. Au cours des dernières années, des études épidémiologiques basées sur des outils de diagnostic améliorés ont estimé que la prévalence réelle des infections à S. stercoralis est beaucoup plus élevée que ce qui avait été rapporté précédemment, soit environ 610 millions de personnes dans le monde2.
S. stercoralis et d’autres espèces de Strongyloides, y compris le parasite de rat étroitement apparenté et le modèle de laboratoire commun S. ratti, ont un cycle de vie inhabituel qui est avantageux pour les études génomiques expérimentales car il se compose à la fois de générations parasites et libres (environnementales)3 (Figure 1). Plus précisément, S. stercoralis et S. ratti peuvent traverser une seule génération de vie libre. La génération de la vie libre se compose de larves post-parasites qui se développent en mâles et femelles adultes libres; toutes les descendants des adultes libres se développent en larves infectieuses, qui doivent infecter un hôte pour poursuivre le cycle de vie. De plus, cette génération environnementale ou libre peut être manipulée expérimentalement en laboratoire. Étant donné que les adultes Strongyloides vivant en liberté et les adultes de C. elegans partagent une morphologie similaire, des techniques telles que la microinjection intragonadale qui ont été développées à l’origine pour C. elegans peuvent être adaptées pour être utilisées avec des Strongyloides 4,5 adultes vivant en liberté. Alors que l’ADN est généralement introduit chez les femelles adultes libres, les mâles et les femelles de Strongyloides peuvent être microinjectés6. Ainsi, des outils génomiques fonctionnels sont disponibles pour interroger de nombreux aspects de la biologie des Strongyloides. D’autres nématodes parasites n’ont pas de génération libre et, par conséquent, ne se prêtent pas aussi facilement aux techniques génomiques fonctionnelles3.
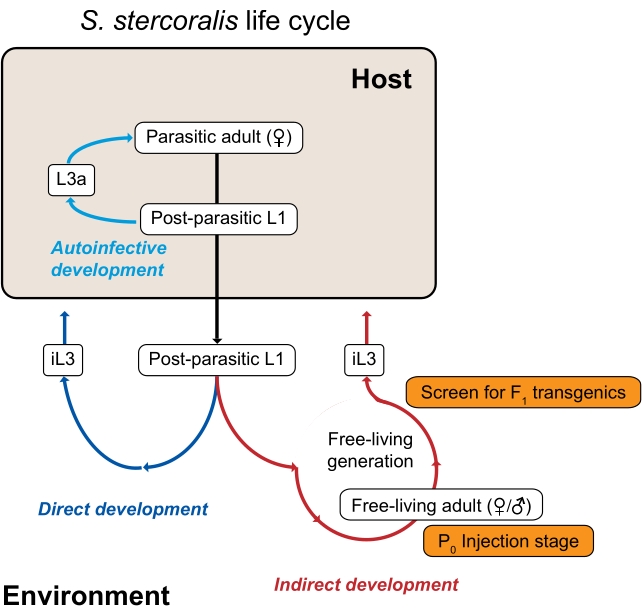
Figure 1 : Cycle de vie de Strongyloides stercoralis. Les femelles parasites de S. stercoralis habitent l’intestin grêle de leurs hôtes mammifères (humains, primates non humains, chiens). Les femelles parasites se reproduisent par parthénogenèse et pondent des œufs dans l’intestin grêle. Les œufs éclosent encore à l’intérieur de l’hôte en larves post-parasites, qui sont ensuite passées dans l’environnement avec des matières fécales. Si les larves post-parasites sont des mâles, elles se développent en mâles adultes libres. Si les larves postparasitaires sont des femelles, elles peuvent se développer en femelles adultes libres (développement indirect) ou en larves infectieuses de troisième stade (iL3s; développement direct). Les mâles et les femelles libres se reproduisent sexuellement pour créer une progéniture qui est contrainte de devenir des iL3. Dans certaines conditions, S. stercoralis peut également subir une auto-infection, dans laquelle certaines des larves postparasitaires restent à l’intérieur de l’intestin hôte plutôt que de passer dans l’environnement dans les matières fécales. Ces larves peuvent se développer en larves auto-infectieuses (L3a) à l’intérieur de l’hôte, pénétrer à travers la paroi intestinale, migrer à travers le corps et éventuellement retourner dans l’intestin pour devenir des adultes reproducteurs. Le cycle de vie de S. ratti est similaire, sauf que S. ratti infecte les rats et n’a pas de cycle auto-infectieux. La génération environnementale est essentielle à l’utilisation des espèces de Strongyloides pour les études génétiques. Les femelles adultes libres (P0) peuvent être microinjectées; leur progéniture, qui deviendront toutes des iL3, sont les transgéniques potentiels F1. Ce chiffre a été modifié à partir de Castelletto et al. 3. Veuillez cliquer ici pour voir une version agrandie de cette figure.
S. stercoralis partage de nombreux aspects de sa biologie avec d’autres nématodes gastro-intestinaux parasitaires humains, y compris l’invasion de l’hôte et la modulation immunitaire de l’hôte. Par exemple, les ankylostomes parasitaires humains des genres Necator et Ancylostoma infectent également par pénétration cutanée, naviguent de manière similaire dans le corps et résident finalement comme des adultes parasites dans l’intestin grêle7. Ainsi, de nombreux nématodes gastro-intestinaux utilisent probablement des comportements sensoriels courants et des techniques d’évasion immunitaire. En conséquence, les connaissances glanées auprès de Strongyloides compléteront les découvertes chez d’autres nématodes moins génétiquement traitables et conduiront à une compréhension plus complète de ces parasites complexes et importants.
Ce protocole de micro-injection décrit la méthode d’introduction de l’ADN dans les femelles adultes strongyloïdes vivant librement pour fabriquer une progéniture transgénique et mutante. Les exigences en matière d’entretien de la souche, y compris le calendrier de développement des vers adultes pour les microinjections et la collecte de la progéniture transgénique, sont décrites. Des protocoles et une démonstration de la technique complète de micro-injection, ainsi que des protocoles pour la culture et le dépistage de la progéniture transgénique, sont inclus, ainsi qu’une liste de tous les équipements et consommables nécessaires.
