Sinds Salvador Moncada en Nobelprijswinnaars Robert Furchgott, Louis Ignarro en Ferid Murad stikstofmonoxide (NO) identificeerden als de eerder bekende endotheel-afgeleide relaxatiefactor (EDRF), is de centrale rol van NO vastgesteld in verschillende belangrijke mechanismen die zich uitstrekken over vasculaire biologie, neurowetenschappen, metabolisme en gastheerrespons 1,2,3,4,5,6,7 . Exogene toediening van NO-gas is een gevestigde behandeling geworden voor respiratoire insufficiëntie als gevolg van pulmonale hypertensie bij de pasgeborene8. Stikstofmonoxidegas is ook onderzocht voor de behandeling van respiratoir syncytieel virus (RSV) infectie, malaria en andere infectieuze ziekten, ischemie-reperfusieletsel en voor de preventie van acuut nierletsel bij patiënten die hartchirurgie ondergaan 9,10,11,12. De behoefte aan nauwkeurige meettechnieken om de niveaus van NO, de metabolieten en die van de doeleiwitten en -verbindingen te beoordelen, komt voort uit zowel mechanistische als interventionele studies.
Vanwege de hoge reactiviteit kan NO verschillende reacties ondergaan, afhankelijk van de biologische matrix waarin het wordt geproduceerd en / of vrijgegeven. Bij afwezigheid van hemoglobine (Hb) of andere oxy-hemoproteïnen wordt NO bijna volledig geoxideerd tot nitriet (NO2–).
2NO + O2 → 2NO2
NO2 + NO → N2O3
N2O3 + H2O → NO2– + H+
NO ondergaat eerst autoxidatie met moleculaire zuurstof (O2) om stikstofdioxide (NO2) te verkrijgen en reageert met NO2 zelf om dinitrogentrioxide te genereren (N2O3). Eén molecuul van N2O3 reageert met water (H2O) om twee moleculen van NO2– en een proton (H+)13 te vormen. In volbloed worden14,15, NO en NO2– snel omgezet in nitraat (NO3–) omdat deze moleculen gretig reageren met de geoxideerde heemgroepen van Hb [Hb-Fe2+-O2 of oxyhemoglobine (oxyHb)] tot NO3–. Deze reactie gaat gepaard met de overgang van de heemgroep naar de ijzertoestand [Hb-Fe3+ of methemoglobine (metHb)]:
Hb-Fe2+-O2 + NO → Hb-Fe3+ + NO3–
De barrière van de rode bloedcel (RBC) en de ruimte direct grenzend aan het endotheel zijn de belangrijkste factoren die deze reactie beperken en waardoor een klein deel van het NO dat door het endotheel vrijkomt, kan fungeren als EDRF16,17. In feite is bekend dat celvrij Hb in de bloedsomloop de vaatverwijding in experimentele en klinische omgevingen verstoort17,18. Binnen de RBC reageert, afhankelijk van oxygenatie en NO 2-concentratie, een deel van NO met deoxyhemoglobine (Hb-Fe2+) om ijzer-nitrosyl Hb (Hb-Fe2+-NO of HbNO) te vormen:
Hb-Fe2+ + NO → Hb-Fe2+-NO
In de RBC15,17 kan NO2– Hb-Fe3+ vormen door Hb-Fe2+ te verlagen wat leidt tot het vrijkomen van NO, wat op zijn beurt Hb-Fe2+-O 2 (bij voorkeur) of Hb-Fe2+ bindt.
De generatie van NO-derivaten moet niet strikt unidirectioneel worden beschouwd, aangezien NO kan worden geregenereerd uit NO2– en NO3– in verschillende weefsels en door verschillende enzymen (bijvoorbeeld door darmbacteriën of in mitochondriën, met name onder hypoxische omstandigheden)19,20.
Een variabele hoeveelheid geproduceerd (of toegediend) NO leidt tot de stroomafwaartse generatie van S-nitrosothiolen, voornamelijk door thioltransnitrosatie van N2O3 in aanwezigheid van een nucleofiel die een NO+ donortussenproduct creëert (Nuc-NO+-NO2–):
N2O3 + RS– → RS-NO + NO2–
Een andere mogelijkheid voor het genereren van S-nitrosothiolen is nitrosylering van geoxideerde thiolen (NO reageren met een geoxideerd thiol):
RS• + NO → RS-NO
Dit mechanisme en de directe thioloxidatie met NO2 zouden alleen mogelijk kunnen zijn in zeer specifieke omstandigheden die elders worden beschreven21. S-nitrosothiolen variëren van lichte moleculen zoals S-nitrosoglutathione tot grote thiolbevattende eiwitten. S-nitrosohemoglobine (S-NO-Hb) wordt gevormd door nitrosatie van een thiolgroep van een geconserveerd cysteïneresidu in de β-keten (β93C)22.
De generatie en het metabolisme van S-nitrosothiolen maken deel uit van belangrijke regulerende mechanismen. Voorbeelden hiervan zijn regulatie van glutathion, caspasen, N-methyl-D-Aspartaat (NMDA) en ryanodinereceptoren 23,24,25,26,27,28. Eerder beschouwd als een belangrijke mediator van NO-biologie in vivo, lijkt nitrosated albumine (S-nitroso-albumine) een NO / NO + transporter te zijn zonder enige specifieke extra biologische activiteit29.
Bij het meten van de concentratie van NO en zijn derivaten van een specifiek biologisch monster binnen een biologische matrix, is het belangrijk om rekening te houden met kenmerken zoals zuurgraad, oxygenatie, temperatuur en de aanwezigheid van reagentia. Voorbeelden hiervan zijn toegediende exogene NO-donoren en, in de setting van acute ontsteking, waterstofperoxide (H2O2) dat reageert met NO2 , wat leidt tot het genereren van supernormale concentratie van vrije radicalen zoals peroxynitriet (ONOO–)21. Naast de analysemethode die wordt gebruikt, is de preanalytische fase van monstervoorbereiding en -opslag van fundamenteel belang. Stroomafwaartse reacties die niet de in vivo NO-activiteit vertegenwoordigen, moeten worden voorspeld, overwogen en geblokkeerd. Een geldig voorbeeld is de instabiliteit van S-NO-Hb, die een speciale behandeling van bloedmonsters vereist wanneer deze is gericht op meting22.
Chemiluminescentie-gebaseerde assays zijn de gouden standaard voor het detecteren van de niveaus van NO en zijn belangrijkste metabolieten [NO2–, NO3–, S-NO en ijzer-nitrosylcomplexen (Fe-NO)] in elke biologische vloeistof, inclusief weefselhomogenaten30,31. Deze methoden zijn gebaseerd op de chemiluminescentiedetector (CLD), een apparaat dat de reactie van NO met ozon (O3) herbergt en NO2 genereert in een aangeslagen toestand (NO2•). Relaxatie van NO2• veroorzaakt emissie van een foton van licht dat wordt gedetecteerd door een fotomultiplicatorbuis, waardoor een elektrisch signaal wordt gegenereerd dat recht evenredig is met het NO-gehalte van het bemonsterde gasmengsel32. Een vereenvoudigd schema van de CLD wordt weergegeven.
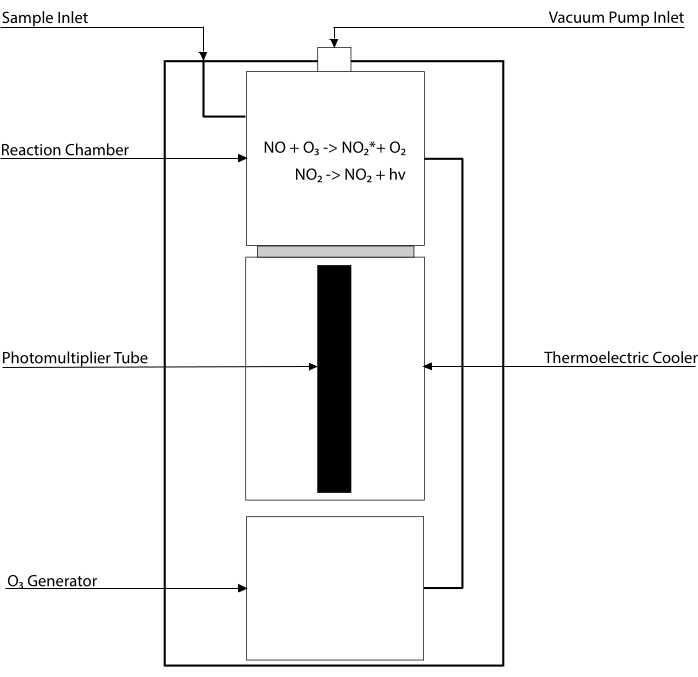
Figuur 1: Vereenvoudigd schema van een chemiluminescentiedetector voor stikstofmonoxidegas. Chemiluminescentie-gebaseerde detectie van stikstofmonoxide (NO) is de stoichiometrische generatie van één foton per NO-gasmolecuul die wordt geïntroduceerd in de chemiluminescentiedetector (CLD). De chemiluminescentiereactie wordt verkregen in een aangewezen kamer die wordt geleverd met ozon (O3) van een interne generator, die op negatieve druk wordt gehouden door verbinding met een externe pomp, waardoor een continue en constante instroom van monstergas mogelijk is. De generatie van O3 vereist diatomische zuurstof (O2) die wordt geleverd door een speciale O2-tank die is aangesloten op de CLD (andere fabrikanten leveren CLD’s die werken met omgevingslucht). In de reactiekamer reageert elk molecuul NO-gas in het bemonsterde gas met zuurstof om één molecuul stikstofdioxide in de geactiveerde toestand (NO2*) op te leveren. Door terug te keren naar zijn grondtoestand zendt elk NO2*-molecuul één foton uit dat wordt gedetecteerd door een fotomultiplicatorbuis (PMT) die zich naast de reactiekamer bevindt. De PMT met de bijbehorende versterker en centrale verwerkingseenheid produceert een signaal dat evenredig is met het aantal fotonen en het aantal NO-moleculen in de reactiekamer. Klik hier om een grotere versie van deze figuur te bekijken.
De monsterinlaat van de CLD kan worden aangesloten op een glaswerksysteem met een reactiekamer voor vloeistofmonsters. Het systeem wordt continu gezuiverd met een inert gas zoals stikstof, helium of argon, waardoor NO van de reactiekamer naar de CLD wordt overgebracht. Vloeistoffasemonsters worden via een speciaal membraan in het spoelvat geïnjecteerd.
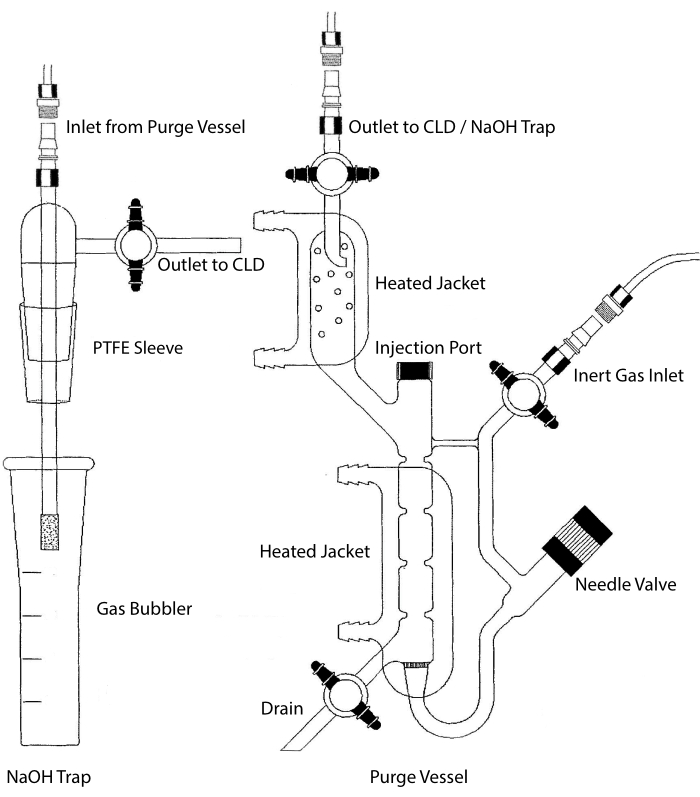
Figuur 2: Structuur van een zuiveringsvat voor op chemiluminescentie gebaseerde detectie van stikstofmonoxidegas Het spoelvat (rechts) maakt de detectie van stikstofmonoxide (NO) gas of een andere verbinding mogelijk die gemakkelijk kan worden omgezet in NO-gas wanneer deze vrijkomt uit een vloeibaar fasereagens. De inerte gasinlaat is verbonden met een bron (tank) van een inert gas zoals Argon, Xeon of diatomische stikstof (N2). De naaldklep (opent aan de linkerkant) wordt gebruikt voor drukregeling in het spoelvat en kan volledig worden verwijderd om het vat te reinigen. De injectiepoort wordt afgedekt door een dop met een membraantussenschot voor monsterinjectie. Het membraan moet vaak worden vervangen. Een verwarmde mantel omringt de reactiekamer en is aangesloten op een warmwaterbad om de VCl3 in HCl-assay uit te voeren. De afvoer van het spoelvat is aangesloten op de chemiluminescentiedetector (CLD) of op de NaOH-val (vereist voor VCl3 in HCl-assays). Om de inhoud van de reactiekamer af te tappen, sluit u eerst de stopkranen bij de inerte gasinlaat en de afvoer van het spoelvat, sluit u de naaldklep, verwijdert u de dop bij de injectiepoort en opent u ten slotte de stopkraan bij de afvoer. De NaOH-val (links) moet in lijn tussen het spoelvat en de CLD worden geplaatst als de VCl3 in HCl-test wordt uitgevoerd vanwege de corrosie van HCl. De verbinding met de CLD vereist altijd dat een intens veld diëlektrisch (IFD) filter wordt geplaatst tussen de CLD en de uitgang van het spoelvat (of de NaOH-val, indien gebruikt). Het IFD-filter verwijdert deeltjes in de lucht en voorkomt dat vloeistof door het spoelvat gaat. PTFE = polytetrafluorethyleen. Klik hier om een grotere versie van deze figuur te bekijken.
Als gevolg hiervan kan elke verbinding die door een specifieke en gecontroleerde chemische reactie in NO kan worden omgezet, met hoge gevoeligheid worden gedetecteerd in elke biologische vloeistof en weefselhomogenaat24. Directe meting van gasfase NO door chemiluminescentie wordt uitgevoerd in zowel experimentele als klinische omgevingen. Deze technieken worden elders uitgebreid beschreven 33,34,35. Meting van NO2–, S-nitrosothiolen, S-nitrosated eiwitten en Fe-NOs kan worden uitgevoerd door monsters toe te voegen in een reactiemengsel met triiodide (I3–), dat stoichiometrisch GEEN gas vrijgeeft uit al deze verbindingen:
I3– → I2 + I–
2NO2− +2I− +4H+ → 2NO + I2 +2H2O
I3− + 2RS-NO → 3I− + RSSR + 2NO+
2NO+ + 2I− → 2NO + I2
terwijl ik3– niet reageert met NO 3-15. Nauwkeurige metingen van elke verbinding worden mogelijk gemaakt door voorbehandeling van monsters met aangezuurd sulfanilamide (AS) met of zonder kwikchloride (HgCl2). In het bijzonder verwijdert de voorbehandeling met AS alle NO 2-inhoud. Als gevolg hiervan weerspiegelt het NO-gehalte gemeten door de CLD alleen de som van de concentratie van S-NOs en Fe-NOs. Injectie van HgCl2 in een monster aliquot vóór AS-injectie zorgt ervoor dat NO2– wordt vrijgegeven door S-NO. Behandeling met AS (leidend tot NO 2-verwijdering) zorgt ervoor dat het gemeten NO-gehalte alleen de concentratie van Fe-NOs weerspiegelt. Een reeks aftrekkingen tussen de beoordelingen maakt het mogelijk om de precieze concentratie van de drie NO-derivaten te berekenen22.
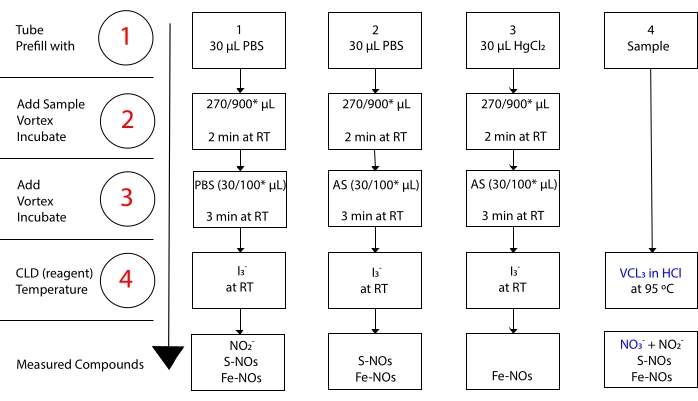
Figuur 3: Stappen in monstervoorbereiding voor de I3– in azijnzuur chemiluminescentietest. De opeenvolgende stappen voor de bereiding van de I3– in azijnzuur chemiluminescentietest worden geïllustreerd. Het gebruik van lichtbeveiligde centrifugebuizen is vereist. De buizen 1, 2 en 3 worden gebruikt om de test voor te bereiden. Een ander monster aliquot (buis 4) is nodig voor de VCl3 in HCl-test als de meting van nitraat (NO3–) vereist is. Stappen worden aangegeven met cijfers in het rood. Voorvullen (stap 1) zoals aangegeven met fosfaatbufferzoutoplossing (PBS) of HgCl2 voordat het monstervolume wordt toegevoegd. Voeg het monstervolume (2) toe zoals aangegeven, vortex, en incubeer gedurende 2 minuten bij kamertemperatuur (RT). Voeg (3) PBS of aangezuurd sulfanilamide (AS) toe zoals aangegeven, vortex en incubeer gedurende 3 minuten bij RT. Voer de test uit (4). De concentratie gemeten met de test is de som van de concentratie van de verbindingen die onder elke buis worden gerapporteerd. Buis nummer 1 maakt metingen van nitriet (NO2–), S-nitrosothiolen (S-NO) en ijzer-nitrosylcomplexen (Fe-NOs) als één signaal mogelijk. Voor nitraatmetingen (NO3-) moeten monsters worden uitgevoerd met zowel I3– in azijnzuur als VCl3 in HCl-assays, en de waarde verkregen uit buis 1 moet worden afgetrokken van die verkregen uit buis 4. *voorgestelde hoeveelheden die moeten worden gebruikt voor Hb-analyse voor de bepaling van restnummer2–, S-nitrosohemoglobine en ijzer-nitrosyl-hemoglobine. Klik hier om een grotere versie van deze figuur te bekijken.
Voor NO 3-meting wordt Vanadium (III) chloride (VCl3) in zoutzuur (HCl) gebruikt voor de omzetting van NO3– in NO in het spoelvat om NO3– stoichiometrisch te meten met de CLD:
2 NO3–+ 3V+3 + 2H2O → 2NO + 3VO2+ + 4H+
Om een voldoende snelle omzetting te bereiken, moet de reactie worden uitgevoerd bij 90-95 °C. Reductie van NO3– naar NO2– gaat gepaard met reductie van NO2– naar NO door HCl. Vanadiummetaal vermindert ook S-NOs waardoor hun NO-deelwordt bevrijd 22,36. De eindconcentratie verkregen door CLD met VCl3 in HCl weerspiegelt de totale concentratie van NO3–, NO2 en andere nitrosaatverbindingen. Aftrekking van deze laatste waarde van de met CLD met I3– opgewekte concentratie maakt het mogelijk om NO3– concentratie36,37 te berekenen (figuur 3).
In de NO-consumptietest genereert de continue afgifte van NO in het spoelvat door NO-donoren zoals (Z)-1-[2-(2-aminoethyl)-N-(2-ammonioethyl)amino]diazen-1-ium-1,2-diolaat (DETA-NONOaat) een stabiel signaal waardoor celvrije oxyHb in de geïnjecteerde monsters kan worden gekwantificeerd. De hoeveelheid NO die in het zuiveringsvat wordt verbruikt, staat in een stoichiometrische relatie met de hoeveelheid oxyHb in het monster38.
Protocollen voor het meten van NO2–, NO3–, S-nitrosothiolen, ijzer-nitrosylcomplexen en NO-consumptie door celvrij Hb in plasmamonsters worden geïllustreerd. Studies naar NO in de RBC-omgeving vereisen een specifieke monsterbehandeling gevolgd door exclusiechromatografie om extreem fragiele S-NO-Hb en Hb-NO te meten in combinatie met de bepaling van de totale Hb-concentratie15,22. Monstervoorbereiding is instrumenteel bij het corrigeren van metingen. Het pre-bestaan van NO2– in H2O en het vrijkomen van NO2– tijdens de test kan leiden tot meting van kunstmatig hogere concentraties NO-derivaten zoals S-NO-Hb14,39. Belangrijke aspecten van monstervoorbereiding worden ook gepresenteerd.
