إن فهمنا لتمايز الخلايا ونشأة الأنسجة والأعضاء هو نتيجة عقود من الشاشات المستهدفة المعقدة للجينات ومنتجاتها. إن زيادة معرفتنا بجميع الجزيئات الحيوية وكمياتها خلال الأحداث الخلوية المهمة من شأنه أن يساعد في كشف الآليات الجزيئية التي تتحكم في الأنماط المكانية والزمانية لخطة جسم الفقاريات. أصبحت التقنيات التي تمكن من التضخيم الجزيئي والتسلسل قادرة الآن على الإبلاغ بشكل روتيني عن أعداد كبيرة من الجينات والنسخ ، مما يدعم الدراسات القائمة على الفرضيات في البحوث البيولوجية الأساسية والانتقالية. لفهم الأنظمة النامية ، تدعو العلاقة المعقدة بين النسخ والترجمة إلى التحليل المباشر للبروتينات المتعددة وتعديلاتها اللاحقة للترجمة. بدأت البروتينات العالمية التي تستخدم الأنظمة البيولوجية في المختبر ، مثل الخلايا الجذعية المستحثة متعددة القدرات ، في تحديد آليات تحريض الأنسجة 1,2. في الكائنات الحية المعقدة ، مثل جنين الفقاريات ، يعتمد التطور على تدرجات المورفوجين في سياق المكان والزمان3. ويترتب على ذلك أن اكتساب المعرفة بالتغيرات البروتينية مع تمايز الخلايا لتشكيل أنسجة متخصصة ، مثل الأنسجة العصبية ، يوفر مفتاحا لإطلاق البرامج الجزيئية التي تتحكم في التطور الطبيعي والمعيب وتوجيه علاجات الجيل التالي.
الضفدع ذو المخالب الفقارية في جنوب إفريقيا (Xenopus laevis) هو نموذج راسخ في البيولوجيا الخلوية والتنموية والعصبية والتجديدية. سلطت جائزة نوبل في علم وظائف الأعضاءأو الطب 4,5 لعام 2012 التي حصل عليها السير جون جوردون لاكتشاف تعدد قدرات النواة الجسدية الضوء على أهمية هذا النموذج للاكتشافات في الدراسات الأساسية والانتقالية. تتطور أجنة Xenopus خارجيا إلى الأم ، مما يسهل التلاعب المباشر بالخلايا واستنساخ الخلايا والتعبير الجيني على مراحل مختلفة من التطور. مكن التصبغ غير المتماثل وانقسامات الخلايا النمطية من رسم خرائط المصير القابلة للتكرار من الجنين المكون من 16-6 و 32 خلية 7,8. بالنسبة للبروتينات القائمة على قياس الطيف الكتلي عالي الدقة (HRMS) ، تشمل المزايا الإضافية للنموذج حجما كبيرا نسبيا (~ 1 مم في القطر) ، والذي ينتج محتوى بروتينيا وفيرا للتحليل (~ 130 ميكروغرام في أجنة مرحلة الانقسام المبكرة ، ~ 10 ميكروغرام من محتوى البروتين في خلايا مفردة من الجنين المكون من 16 خلية)9,10.
في الوقت الحاضر ، HRMS هي التكنولوجيا الرائدة المفضلة للكشف عن البروتينات. تتيح هذه التقنية الكشف المباشر والحساس والمحدد والقياس الكمي للبروتينات المتعددة ، وعادة ما تكون مئات إلى آلاف البروتينات المختلفة11. تتضمن البروتينات من أسفل إلى أعلى بواسطة HRMS سلسلة من الخطوات المترابطة. بعد الاستخراج من عينة الخلية / الأنسجة ، يتم هضم البروتينات بإنزيم محلل للبروتين ، مثل التربسين (البروتينات من أسفل إلى أعلى). يتم فصل الببتيدات الناتجة بناء على خصائصها الفيزيائية والكيميائية المختلفة ، بما في ذلك الكارهة للماء (كروماتوغرافيا السائل ذات الطور العكسي ، LC) ، الشحنة الصافية (كروماتوغرافيا التبادل الأيوني) ، الحجم (كروماتوغرافيا استبعاد الحجم) ، أو الحركة الكهربائية (الرحلان الكهربائي الشعري ، CE). ثم يتم شحن الببتيدات (المتأينة) ، عادة باستخدام التأين بالرش الكهربائي (ESI) ، ويتم الكشف عن أيونات الببتيد وتسلسلها عبر تجزئة الطور الغازي بواسطة نظام إدارة الموارد البشرية الترادفي. يتم تعيين بيانات الببتيد الناتجة إلى بروتين الكائن الحي قيد الدراسة. مع ارتباط شدة إشارة أيون الببتيد الخاصة بالبروتين (البروتيني) بالتركيز ، يمكن إجراء القياس الكمي للبروتين بدون ملصق أو قائم على الملصق (تكميم تعدد الإرسال). تنتج بروتينات HRMS موردا غنيا من المعلومات حول الحالة الجزيئية للنظام قيد الدراسة ، مما يسمح بتوليد الفرضيات ومتابعة الدراسات الوظيفية.
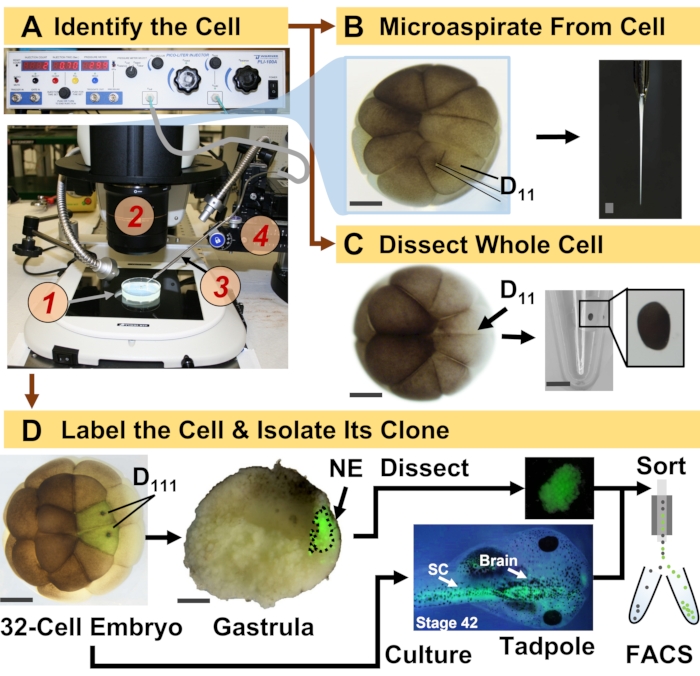
الشكل 1: البروتينات القابلة للتطوير المكاني الزماني التي تمكن بروتينات HRMS الموجهة بنسب الخلية في الجنين النامي (الضفدع). (أ) تصور العينة (1) باستخدام مجهر مجسم (2) لحقن خلية محددة (أقحم) ، باستخدام ماصة دقيقة مصنعة (3) تحت السيطرة من خلال مرحلة الترجمة (4). ب: أخذ عينات تحت خلوية من الخلية D 11 اليسرى المحددة في جنين مكون من16 خلية. ج: تشريح خلية D 11 كاملة من جنين مكون من16 خلية. (د) تتبع الفلورسنت (الأخضر) لنسل D111 الأيسر والأيمن من جنين مكون من 32 خلية لتوجيه تشريح الأديم الظاهر العصبي (NE) في المعدة (المرحلة 10) وعزل النسيج المنحدر من الشرغوف باستخدام FACS. قضبان المقياس: 200 ميكرومتر للأجنة ، 1.25 مم للقارورة. تم تكييف الأرقام بإذن من المراجع15،19،21،59. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
يتيح البروتوكول المقدم هنا القياس الكمي القائم على نظام إدارة الموارد البشرية لأعداد كبيرة من البروتينات في الخلايا / الأنسجة المحددة في أجنة X. laevis النامية. يعتمد النهج على التحديد الدقيق للخلايا ، وخرائط مصير الخلية القابلة للتكرار ، والمنهجيات المعمول بها لتتبع سلالات الخلايا في هذا النموذج البيولوجي6،7،8. كما هو موضح في الشكل 1، ندرس البروتينات من الخلايا المفردة عن طريق استخدام تشريح الخلية الكاملة أو أخذ العينات المجهرية الشعرية لشفط المحتوى الخلوي. تسمح لنا مراقبة نسب الخلية بدراسة التطور الزماني المكاني للبروتين حيث تشكل الخلايا الأنسجة أثناء المعدة. يتم تمييز ذرية الخلية بالفلورسنت عن طريق حقن فلوروفور مترافق مع ديكستران خامل أو mRNA للبروتين الفلوري (على سبيل المثال ، بروتين الفلورسنت الأخضر ، أو GFP). يتم عزل النسل المسمى في النقاط الزمنية التطورية المطلوبة. أثناء المعدة ، يمكن عزل الحيوانات المستنسخة الخلوية المتجمعة بإحكام عن طريق التشريح. بعد المعدة ، يمكن توزيع استنساخ الخلايا داخل الجنين بسبب حركات الهجرة ويمكن عزلها عن الأنسجة المنفصلة عن طريق فرز الخلايا المنشطة بالفلورة (FACS). يتم قياس البروتينات في هذه الخلايا والأنسجة عن طريق البروتينات من أسفل إلى أعلى باستخدام HPLC أو CE للفصل و ESI جنبا إلى جنب HRMS لتحديد الهوية. بروتينات HRMS الموجهة بنسب الخلايا قابلة للتطوير لأحجام وسلالات مختلفة داخل الجنين وهي محددة وحساسة وكميتة. من خلال أمثلة مختارة موضحة هنا ، نوضح أيضا أن هذا البروتوكول قابل للتطوير وقابل للتكيف على نطاق واسع مع أنواع مختلفة من الخلايا وسلالات الخلايا.
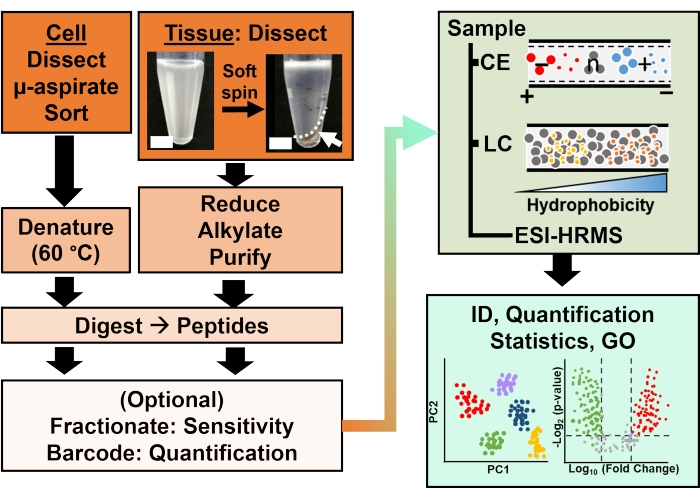
الشكل 2: سير عمل التحليل الحيوي. سهل التشريح الدقيق والشفط الشعري ، أو FACS أخذ عينات من محتوى البروتين الخلوي والنسيلي. استنفاد بروتينات صفار البيض الوفيرة وفصلها عن طريق الرحلان الكهربائي الشعري (CE) أو كروماتوغرافيا السائل ذات التدفق النانوي (LC) حساسية التعريف المعززة (ID) باستخدام التأين الكهربائي (ESI) قياس الطيف الكتلي عالي الدقة (HRMS). كشف القياس الكمي عن عدم التنظيم ، وتوفير معلومات جديدة للدراسات القائمة على الفرضيات بالتزامن مع المعلومات المتاحة من علم الوجود الجيني (GO). تم تكييف الأرقام بإذن من المرجع15. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
