En fælles karakteristika for lidelser og sygdomme i centralnervesystemet (CNS), såsom traumatisk hjerneskade (TBI), rygmarvsskade (SCI), slagtilfælde, Alzheimers sygdom og Parkinsons sygdom er afbrydelsen af aksonale veje og neuronale celler Tab 1 , 2 , 3 , 4 , 5 , 6 . For eksempel, når et iskæmisk slagtilfælde går ubehandlet, estimeres det, at axoner går tabt med en hastighed på 7 miles af axoner pr. Minut 5 . I tilfælde af TBI, som ca. 1,7 millioner mennesker oplever hvert år alene i USA, kan axonal degeneration fortsætte med at forekomme år efter traumer, da den oprindelige skade udfælder en langtids neurodegenerativ tilstand 4 . Forværring af disse skadelige virkninger har CNS en stærkt begrænset capaBy til regenerering 1 , 7 , 8 , 9 . Efter skader udvikler et hæmmende miljø, der er kendetegnet ved manglende rettet vejledning til fjerne mål, tilstedeværelsen af myelin-associerede inhibitorer, der forhindrer neuritudvæksten og dannelsen af et glialær ved reaktive astrocytter 8 , 10 , 11 , 12 . Glialæren tjener som en biokemisk og fysisk barriere for regenerering med molekyler som chondroitinsulfatproteoglycaner, der forhindrer axonudvækst 8 , 11 . Selvom neurale stamceller er blevet fundet i det voksne CNS, er produktionen af nye neuroner begrænset, da konsistente tegn på neurogenese kun er blevet fundet i lygtepæren, den hippocampaleSubgranulær zone, det periventrikulære område og den centrale kanal i rygmarven 13 , 14 . Disse hindringer forhindrer funktionel genopretning af tabte neuroner og hvid materiel arkitektur efter skade eller sygdom, hvilket resulterer i de ofte livsforandrende og langvarige virkninger af disse tilstande.
På trods af manglen på regenerativ kapacitet i det voksne CNS, er det blevet påvist, at axonal regenerering er mulig, hvis der fremlægges passende miljømærker til værtsneuroner 15 , 16 , 17 , 18 . Forskere har forsøgt at levere og manipulere vækstfaktorer ( fx nervevækstfaktor, epidermal vækstfaktor, glialafhængig vækstfaktor og neurotrofisk faktor-3) og andre vejledningsmolekyler til stimulering af plasticitet og axonregenerering 14 , </ Sup> 18 , 19 . Selv om disse undersøgelser har bekræftet, at voksne axoner er i stand til at reagere på vækstfaktorer, er disse strategier begrænset af den lave permeabilitet af blodhjernebarrieren og de specifikke rumlige og tidsmæssige gradienter, der kræves for at fremme regenerering 14 , 18 , 19 . Andre fremgangsmåder har påberåbt hyperaktiviteten af regenerationsrelaterede transkriptionsfaktorer i CNS neuroner. For eksempel stimulerede overekspression af stat3-transkriptionsfaktoren axonal regenerering i optisk nerve 20 . Ikke desto mindre undlader både biomolekylafgivelse og overekspression af transkriptionsfaktorer at erstatte tabte neuronale populationer. Cellebaserede strategier har hovedsageligt centreret om transplantation af neurale stamceller (NSC'er) i CNS, idet de udnytter deres evne til at erstatte CNS-neuroner, frigive trofiske faktorer,Og støtte de forsøg på neurogenese, der opstår efter skade 17 . På trods af dette er der stadig presserende udfordringer, der forhindrer denne tilgang, herunder transplanterede neurale cellers hæmmede evne til at overleve, integrere med værten og forbliver rumligt begrænset til det skadede område 6 , 14 , 17 , 21 . Desuden er celleafgivelse alene ude af stand til at genetablere cytoarkitekturen af beskadigede eller tabte axonale veje. En alternativ tilgang, der behandler problemerne med celle- og lægemiddel / kemikalieleveringsstrategier, kombinerer disse fremgangsmåder med anvendelsen af biomaterialerne 14 , 22 , 23 . Biomaterialer såsom hydrogeler er i stand til at emulere de biokemiske og fysiske egenskaber af den ekstracellulære matrix (ECM), der hjælper med celletilførsel ogD retention inden for det skadede område og levere vækstfaktorer og andre bioaktive molekyler med kontrolleret frigivelse 22 . De attraktive egenskaber ved disse biomaterialebaserede strategier har resulteret i beviser for in vivo axonal regenerering efter transplantation af stilladser til det læsionerede område 24 , 25 , 26 , 27 , 28 , 29 , 30 . Imidlertid erstatter acellulære biomaterialestrategier ikke tabte neuronale populationer; Når de anvendes som leveringskøretøjer til neuronale, glial- eller neuronale precursorceller, er biomaterialer ikke i stand til at rekonstituere langdistance-axonale netværk. Udfordringen med at udvikle en tilgang, der tackler både den axonale path degeneration og neuronale tab forbundet med CNS skade og sygdom er stadig <Sup class = "xref"> 31.
Vores forskergruppe har tidligere rapporteret udviklingen af implanterbare mikrovæv-manipulerede neurale netværk (mikro-TENN'er), som er en type "levende stillads" bestående af neuronelle celleorganer begrænset til en eller begge ender af en agarosehydrogel-ECM mikrosøjle , Med justerede aksonale kanaler, der strækker sig gennem det indre af denne tredimensionale (3D) indkapsling 1 , 10 , 31 , 32 . En af de største forskelle mellem denne teknik og tidligere tilgange er, at cytoarkitekturen af mikro-TENN'er er skabt helt in vitro og transplanteres bagefter 33 , 34 , 35 , 36 , 37 , 38 , <Sup class = "xref"> 39 , 40 , 41 . In vitro- fremstilling giver omfattende rumlig og tidsmæssig kontrol af cellulær fænotype og orientering, mekaniske / fysiske egenskaber, biokemiske signaler og eksogene faktorer, hvilket gavner værten efter integrationen af disse stilladser efter implantation 41 , 42 . Mikro-tenner er anatomisk inspireret, fordi de efterligner hjernens neuroanatomi, viser axonale kanaler, som ligner dem, der broerer de forskellige funktionelle regioner i hjernen ( figur 1A ) 1 . Derfor kan denne strategi være i stand til fysisk at erstatte tabte hvide stofkerner og neuroner efter implantation i en læsioneret region. Denne teknik er også inspireret af udviklingsmekanismer, hvor "naturlige levende stilladser" dannet af radiale glialceller og banebrydende axoner virker som vejledende guider for celleMigration fra subventriculær zone og aksonal udvækst, henholdsvis 43 . Disse mekanismer er rekapituleret i de justerede aksonale kanaler i mikro-TENN'er, som kan fremvise levende veje for neuralcellemigration og axonal regenerering ved axonformidlet aksonal udvækst ( figur 1C ) 43 . Desuden udnytter denne strategi fordel af synaptisk integration mellem mikro-TENN-neuronerne og det oprindelige kredsløb, der danner nye relæer, der kan bidrage til funktionel opsving ( figur 1B ) 43 . Kapaciteten til synapsdannelse kan også give denne tilgang evnen til at modulere CNS og reagere på værtsvæv ifølge netværksfeedback. For eksempel kan optogenetisk aktive neuroner i de levende stilladser stimuleres til at modulere værtsneuroner gennem synaptiske interaktioner ( figur 1D ).
Derudover vil den biomaterialebaserede rørformede konstrUgn af mikro-tenner giver et passende miljø til celleadhæsion, vækst, neuritilvækst og signalering, mens konstruktionernes miniaturedimensioner muliggør muligvis minimalt invasiv implantering og tilvejebringer en delvis sekvestreret mikromiljø for gradvis integration i hjernen. Faktisk har nyere publikationer vist potentialet hos mikro-tenner til at efterligne neurale veje efter implantation i rottehjerne. Efter stereotaxisk mikroinjektion rapporterede vi tidligere tegn på mikro-TENN-neuronal overlevelse, vedligeholdelse af aksonalkanalarkitektur og neurit forlængelse i værtscortex ud til mindst 1 måned in vivo 10 , 31 . Endvidere tilvejebragte mærkning med synapsin histologisk bevis for synaptisk integration med nativt væv 10 , 31 . Samlet set kan mikro-tennene være særligt velegnede til at rekonstruere og modulere beskadigedeCNS ved at erstatte tabte neuroner, synaptisk integrering med værts kredsløb, genoprette tabt aksonal cytoarkitektur og i visse tilfælde at tilvejebringe regenererende axoner med de relevante vejsøgningsindikatorer.
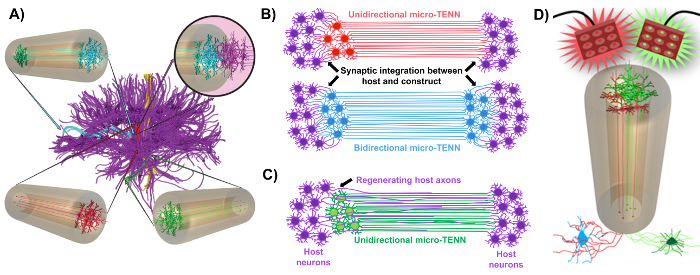
Figur 1: Principper og inspiration bag udviklingen af mikrovæv-manipulerede neurale netværk (mikro-tenner). ( A ) Mikro-TENN'er efterligner cytoarkitekturen i hjernekoblingen (lilla), hvor funktionelt adskilte regioner er forbundet med lange, justerede aksonale kanaler i en retningsretning (rød, grøn) eller tovejs (blå) måde. Som et eksempel kunne mikro-tenner rekonstruere tabte forbindelser i kortikothalamiske og nigrostriatale veje eller i den perforere vej fra entorhinal cortex til hippocampus (tilpasset fra Struzyna et al. , 2015) 1 . ( B ) Diagram af en ensrettet retningL og tovejs mikro-TENN (henholdsvis rød og blå) synaptisk integrering med værts kredsløb (lilla) for at fungere som et funktionelt relæ mellem begge ender af en læsion. ( C ) Skematisk af aksonale kanaler af en ensrettet mikro-TENN (grøn), der tjener som en vejledning til akson-faciliteret regenerering af værtsaksoner (lilla) mod et mål, som mikro-tennene interagerer med. ( D ) Konceptdiagram over brugen af optogenetisk aktive mikro-TENNS som neuromodulatorer, idet man udnytter synaptisk integration med excitatoriske eller hæmmende neuroner (bund). Klik her for at se en større version af denne figur.
Det nuværende manuskript beskriver den metode, der anvendes til fremstilling af mikro-TENN'er ved anvendelse af embryonalt afledte cerebrale kortikale neuroner. Især kan mikro-tenn fremstilles med andre typer af neurale celler. For eksempelDe oprindelige rapporter om succesfuld mikro-TENN-udvikling omfattede dorsale rodganglion (DRG) neuroner 32 . Hydrogelmikrokolonnerne kan genereres ( figur 2A ) ved tilsætning af flydende agarose til et skræddersyet, laserskæret cylindrisk kanalarray eller til kapillarrør, der begge indeholder indstillede akupunkturnåle. Nålen danner lumen og bestemmer mikrokolonnens indvendige diameter (ID), medens kapillarrørets ID og diameteren af cylindrene i den laserskårne anordning dikterer konstruktionernes ydre diameter (OD). OD og ID kan vælges ifølge den ønskede ansøgning ved at vælge forskellige diametre for henholdsvis anordning / kapillærrør og akupunktur nåle. Længden af mikrokolonnerne kan også varieres; Til dato har vi rapporteret opførelsen af mikro-tenner op til 20 mm i længden 10 og arbejder aktivt med længere længder. Efter agarosegelene og akupunkturen nEedler fjernes, en ECM-opløsning, som generelt består af type I-collagen, og laminin tilsættes til konstruktionens lumen ( figur 2C ). ECM-kernen tilvejebringer et stillads til understøttelse af neuronal celleadhæsion og axonal udvækst. I første omgang blev primære rottekortiske neuroner udpladet i mikrokolonnerne under anvendelse af dissocierede cellesuspensioner 10 , 31 , 32 . Denne fremgangsmåde producerede imidlertid ikke mål-cytoarkitekturen i alle tilfælde, som blev defineret som de neuronale cellelegemer, der var begrænset til enderne af mikrokolonnerne, med det centrale lumen bestående af renjusterede aksonale kanaler. Siden da har brugen af en tvungen neuronal aggregeringsmetode (baseret på protokoller tilpasset fra Ungrin et al .) Muliggjort en mere pålidelig og ensartet fremstilling af mikro-tenner med den ideelle struktur ( figur 2B ) 44 . Ud over at beskrive strømmenMetode, vil denne artikel vise repræsentative fasekontrast og konfokale billeder af mikro-TENN'er, der demonstrerer dannelsen af axonale kanaler over tid, såvel som den færdige mål-cytoarkitektur. Dette manuskript vil også udvide på bemærkelsesværdige aspekter af protokollen og de resterende udfordringer og fremtidige retninger af mikro-TENN teknologien.
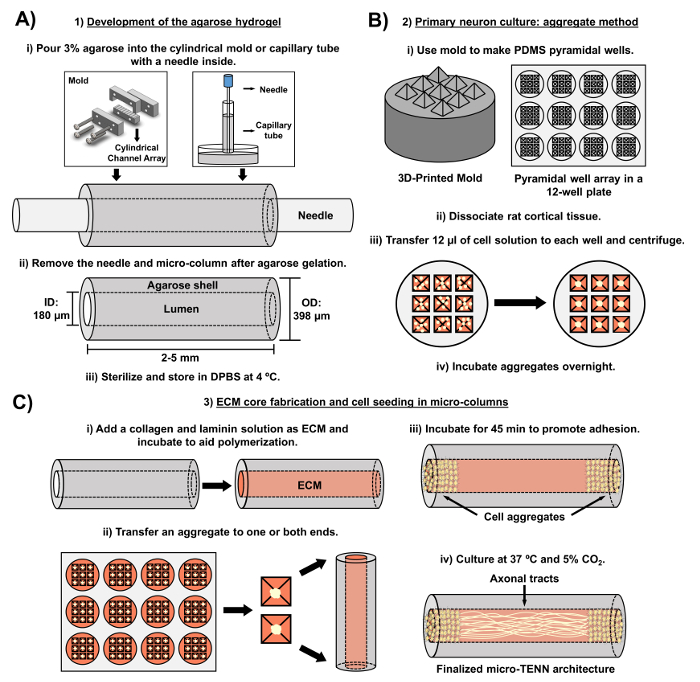
Figur 2: Skematisk diagram over tre-trins mikro-TENN fabrikationsprocessen. ( A ) Udvikling af agarosehydrogelen: (i) Indledningsvis indsættes en lille akupunkturnål ( f.eks . 180-350 μm i diameter) i en cylindrisk kanal af en skræddersyet laserskæret form eller et kapillarrør ( f.eks. , Diameter 380-700 μm). I det næste trin introduceres flydende agarose i DPBS i de cylindriske kanaler eller kapillarrør. (Ii) Efter agarosegelene fjernes nålen ogFormen er adskilt for at give de hule agarosemikolonner. (Iii) Disse konstruktioner steriliseres derefter og opbevares i DPBS. ( B ) Primær neuronkultur og aggregatmetoden: (i) Neuronal aggregering udføres i pyramidale mikrobrøndsarrayer, støbt fra 3D-trykte forme, der passer ind i brøndene på en 12-brønds kulturplade. (Ii) Mikro-TENN'er indbefatter primære rotte neuroner dissocieret fra føtalhjerner af embryonaldag-18 rotter. Efter vævsdissociation med trypsin-EDTA og DNase I fremstilles en celleopløsning med en densitet på 1,0-2,0 x 106 celler / ml. (Iii) 12 μl af denne opløsning overføres til hver brønd i den pyramide mikrobrøndsarray. Pladen indeholdende disse mikrobrønde centrifugeres for at fremstille celleaggregater. (Iv) Disse inkuberes derefter natten over inden plating i mikrokolonnerne. ( C ) ECM-kernefabrikation og celledannelse: (i) Forud for cellesædning blev en ECM-opløsning indeholdende 1 mg / ml type I-kollagen og 1 mg / mlLaminin overføres til det indre af mikro-tennene og får lov til at polymerisere. (Ii) Afhængigt af om ensrettet eller tovejs mikro-tenn fremstilles, anbringes et aggregat i henholdsvis en eller begge ekstremer af mikrokolonnen. (Iii) Efter en inkubationsperiode for at fremme vedhæftning dyrkes mikro-TENN i petriskåle oversvømmet med suppleret embryonalt neuronalt basalt medium. Iv) Efter 3-5 dage i dyrkning skal den endelige mikro-TENN-struktur demonstrere celleaggregater ved yderpunkterne af mikrokolonnen, med aksonale kanaler, der strækker sig over længden. Klik her for at se en større version af denne figur.
