Ein gemeinsames Merkmal von Erkrankungen und Erkrankungen des Zentralnervensystems (ZNS), wie z. B. traumatische Hirnverletzungen (TBI), Rückenmarksverletzungen (SCI), Schlaganfall, Alzheimer-Krankheit und Parkinson-Krankheit, ist die Trennung von axonalen Bahnen und neuronaler Zelle Verlust 1 , 2 , 3 , 4 , 5 , 6 . Zum Beispiel, wenn ein ischämischer Schlaganfall unbehandelt wird, wird geschätzt, dass Axone mit einer Rate von 7 Meilen von Axonen pro Minute 5 verloren gehen. Im Falle von TBI, die etwa 1,7 Millionen Menschen jedes Jahr in den USA alleine erleben, kann die axonale Degeneration auch nach dem Trauma weiter auftreten, da die anfängliche Verletzung einen langfristigen neurodegenerativen Zustand auslöst 4 . Diese schädlichen Wirkungen verschärfen, hat das ZNS eine stark eingeschränkte KapazitätStadt zur Regeneration 1 , 7 , 8 , 9 . Nach der Verletzung entwickelt sich eine hemmende Umgebung, die durch einen Mangel an gerichteter Orientierung an entfernten Targets, die Anwesenheit von Myelin-assoziierten Inhibitoren, die das Auswachsen von Neuriten behindern, und die Bildung einer Glia-Narbe durch reaktive Astrozyten 8 , 10 , 11 , 12 gekennzeichnet ist . Die gliale Narbe dient als biochemische und physikalische Barriere gegen die Regeneration, wobei Moleküle wie Chondroitinsulfat-Proteoglykane das Axonwachstum 8 , 11 behindern. Darüber hinaus, obwohl neuronale Stammzellen in der erwachsenen ZNS gefunden worden sind, ist die Produktion von neuen Neuronen begrenzt, da konsequente Hinweise auf Neurogenese wurde nur in der olfaktorischen Glühbirne, die Hippocampal gefundenSubgranulare Zone, die periventrikuläre Fläche und der zentrale Kanal des Rückenmarks 13 , 14 . Diese Hindernisse verhindern die funktionelle Wiederherstellung von verlorenen Neuronen und der weißen Stoffarchitektur nach Verletzung oder Krankheit, was zu den oft lebensverändernden und verlängerten Wirkungen dieser Bedingungen führt.
Trotz des Mangels an regenerativer Kapazität im adulten ZNS wurde gezeigt, dass eine axonale Regeneration möglich ist, wenn den Wirtsneuronen 15 , 16 , 17 , 18 adäquate Umgebungsmerkmale präsentiert werden. Forscher haben versucht, Wachstumsfaktoren zu liefern und zu manipulieren ( zB Nervenwachstumsfaktor, epidermaler Wachstumsfaktor, glialabhängiger Wachstumsfaktor und neurotrophischer Faktor-3) und andere Leitmoleküle zur Stimulierung von Plastizität und Axonregeneration 14 , </ Sup> 18 , 19 Obwohl diese Studien bestätigt haben, dass erwachsene Axone in der Lage sind, auf Wachstumsfaktoren zu reagieren, sind diese Strategien durch die geringe Permeabilität der Blut-Hirn-Schranke und die spezifischen räumlichen und zeitlichen Gradienten, die zur Förderung der Regeneration 14 , 18 , 19 erforderlich sind, begrenzt. Andere Ansätze haben sich auf die Hyperaktivierung von regenerationsbedingten Transkriptionsfaktoren in ZNS-Neuronen gestützt. Zum Beispiel stimulierte die Überexpression des Stat3-Transkriptionsfaktors die axonale Regeneration im Sehnerv 20 . Trotzdem ersetzen sowohl die Biomolekülabgabe als auch die Überexpression von Transkriptionsfaktoren die verlorenen neuronalen Populationen nicht. Zellbasierte Strategien konzentrierten sich vor allem auf die Verpflanzung von neuralen Stammzellen (NSCs) in das ZNS, wobei sie ihre Fähigkeit nutzen, ZNS-Neuronen zu ersetzen, trophische Faktoren freizugeben,Und unterstützen die Neurogeneseversuche, die nach Verletzung auftreten 17 . Trotzdem gibt es immer noch dringende Herausforderungen, die diesen Ansatz behindern, einschließlich der behinderten Fähigkeit, transplantierte neuronale Zellen zu überleben, mit dem Wirt zu integrieren und räumlich auf den verletzten Bereich 6 , 14 , 17 , 21 zu beschränken. Darüber hinaus ist die Zelllieferung allein nicht in der Lage, die Cytoarchitektur von beschädigten oder verlorenen axonalen Wegen wiederherzustellen. Ein alternativer Ansatz, der die Probleme der Zelle und der Arzneimittel- / Chemikalienabgabestrategien anspricht, kombiniert diese Ansätze mit dem Einsatz von Biomaterialien 14 , 22 , 23 . Biomaterialien wie Hydrogele sind in der Lage, die biochemischen und physikalischen Eigenschaften der extrazellulären Matrix (ECM) zu emulieren, wobei sie in der Zellabgabe undD Retention innerhalb des verletzten Bereichs und liefert Wachstumsfaktoren und andere bioaktive Moleküle mit kontrollierter Freisetzung 22 . Die attraktiven Merkmale dieser biomaterialbasierten Strategien haben zu einer Nachweis der in vivo axonalen Regeneration nach der Transplantation von Gerüsten in den Läsionsbereich 24 , 25 , 26 , 27 , 28 , 29 , 30 geführt . Allerdings ersetzen azelluläre Biomaterial-Strategien nicht verlorene neuronale Populationen; Wenn sie als Lieferfahrzeuge für neuronale, gliale oder neuronale Vorläuferzellen verwendet werden, sind Biomaterialien nicht in der Lage, Fern-Axonal-Netzwerke zu rekonstituieren. Die Herausforderung, einen Ansatz zu entwickeln, der sowohl die axonale Wegdegeneration als auch den neuronalen Verlust, der mit der ZNS-Verletzung und der Krankheit verbunden ist, bekämpft, bleibt noch <Sup class = "xref"> 31.
Unsere Forschungsgruppe berichtete bisher über die Entwicklung von implantierbaren mikrogewebetechnischen neuronalen Netzwerken (Mikro-TENNs), die eine Art von "lebendem Gerüst" sind, bestehend aus neuronalen Zellkörpern, die auf ein oder beide Enden einer Agarose-Hydrogel-ECM-Mikrosäule beschränkt sind Mit ausgerichteten axonalen Traktaten, die sich im gesamten Inneren dieser dreidimensionalen (3D) Umhüllung 1 , 10 , 31 , 32 erstrecken. Einer der Hauptunterschiede zwischen dieser Technik und früheren Ansätzen besteht darin, dass die Cytoarchitektur von Mikro-TENNs vollständig in vitro erzeugt und danach nach 33 , 34 , 35 , 36 , 37 , 38 , <transplantiert wirdSup class = "xref"> 39 , 40 , 41 . In-vitro- Fertigung bietet eine ausgedehnte räumliche und zeitliche Kontrolle von zellulärem Phänotyp und Orientierung, mechanisch-physikalischen Eigenschaften, biochemischen Cues und exogenen Faktoren, die die Integration dieser Gerüste mit dem Wirt nach der Implantation 41 , 42 zugute kommen . Mikro-TENNs sind anatomisch inspiriert, weil sie die Hirnneuroanatomie emulieren und axonale Traktate zeigen, die denen ähnlich sind, die verschiedene funktionale Bereiche des Gehirns überbrücken (Abbildung 1A ) 1 . Daher kann diese Strategie in der Lage sein, die verlorenen weißen Materie Traktate und Neuronen nach der Implantation in eine läsionierte Region physisch zu ersetzen. Diese Technik wird auch von Entwicklungsmechanismen inspiriert, in denen "natürliche lebende Gerüste", die durch radiale Gliazellen und wegweisende Axone gebildet werden, als Wegweiser für Zelle dienenMigration aus der subventrikulären Zone und axonale Auswüchse bzw. 43 . Diese Mechanismen werden in den ausgerichteten axonalen Traktaten von Mikro-TENNs rekapituliert, die lebende Wege für die neuronale Zellmigration und die axonale Regeneration durch axonvermitteltes axonales Auswachsen darstellen können (Abbildung 1C ) 43 . Darüber hinaus nutzt diese Strategie die synaptische Integration zwischen den Mikro-TENN-Neuronen und nativen Schaltkreisen und bildet neue Relais, die zur funktionellen Wiederherstellung beitragen können (Abbildung 1B ) 43 . Die Fähigkeit zur Synapsenbildung kann auch diesem Ansatz die Möglichkeit geben, das ZNS zu modulieren und auf das Gewebe nach Netzwerkrückmeldung zu reagieren. Zum Beispiel können optogenetisch aktive Neuronen in den lebenden Gerüsten stimuliert werden, um Host-Neuronen durch synaptische Wechselwirkungen zu modulieren (Abbildung 1D ).
Darüber hinaus ist die Biomaterial-basierte RohrkonstruktionDie Vermeidung von Mikro-TENNs bietet eine adäquate Umgebung für die Zelladhäsion, das Wachstum, die Neuritenverlängerung und die Signalisierung, während die Miniaturabmessungen der Konstrukte potentiell eine minimal invasive Implantation ermöglichen und eine teilweise sequestrierte Mikroumgebung für eine allmähliche Integration in das Gehirn bieten. In der Tat haben die jüngsten Veröffentlichungen das Potenzial von Mikro-TENNs nachgewiesen, um neuronale Wege nach der Implantation in das Rattenhirn zu imitieren. Nach der stereotaktischen Mikroinjektion haben wir bisher den Nachweis des mikro-TENN-neuronalen Überlebens, die Aufrechterhaltung der axonalen Traktarchitektur und die Neuritenerweiterung in den Wirtskortex mindestens 1 Monat in vivo 10 , 31 gemeldet. Darüber hinaus lieferte die Markierung mit Synapsin histologische Hinweise auf die synaptische Integration mit dem nativen Gewebe 10 , 31 . Insgesamt können Mikro-TENNs eindeutig geeignet sein, um beschädigt zu rekonstruieren und zu modulierenZNS durch Ersetzen verlorener Neuronen, synaptische Integration mit Host-Schaltungen, Wiederherstellung der verlorenen axonalen Cytoarchitektur und in bestimmten Fällen die Bereitstellung von regenerierenden Axonen mit den entsprechenden Pfadfindungs-Cues.
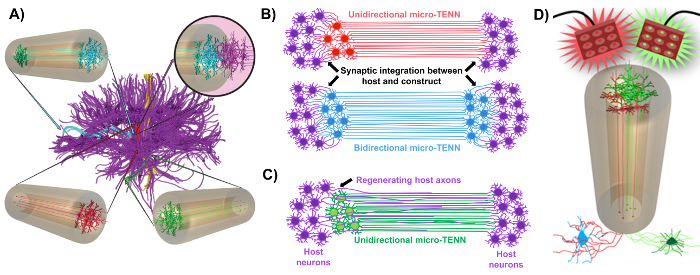
Abbildung 1: Grundsätze und Inspirationen hinter der Entwicklung von mikrogewebetechnischen Neuronennetzen (Mikro-TENNs). ( A ) Micro-TENNs imitieren die Cytoarchitektur des Gehirnverbundes (violett), in der funktionell unterschiedliche Bereiche durch lange, ausgerichtete Axonaltrakte in einer unidirektionalen (rot, grün) oder bidirektionalen (blauen) Weise verbunden sind. Beispielsweise könnten Mikro-TENNs verlorene Verbindungen in kortikothalamischen und nigrostriatalen Bahnen oder im perforanten Weg von der entorhinalen Kortex zum Hippocampus (angepasst von Struzyna et al. , 2015) rekonstituieren. ( B ) Diagramm einer UnidirektionaL und bidirektionale Mikro-TENN (rot und blau) synaptisch mit der Host-Schaltkreise (lila) integriert, um als funktionales Relais zwischen beiden Enden einer Läsion zu dienen. ( C ) Schematische Darstellung der axonalen Traktate eines unidirektionalen Mikro-TENN (grün), die als Leitfaden für die axon-erleichterte Regeneration von Wirts-Axonen (violett) zu einem Ziel dienen, mit dem das Mikro-TENN wechselwirkt. ( D ) Konzeptionelle Darstellung der Verwendung von optogenetisch aktiven Mikro-TENNS als Neuromodulatoren unter Ausnutzung der synaptischen Integration mit exzitatorischen oder inhibitorischen Neuronen (unten). Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
Das aktuelle Manuskript beschreibt die Methodik, die zur Herstellung von Mikro-TENNs unter Verwendung von embryonal abgeleiteten zerebralen kortikalen Neuronen verwendet wird. Bemerkenswerterweise könnten Mikro-TENNs mit anderen Arten von neuralen Zellen hergestellt werden. Für exReichlich, die ersten Berichte über die erfolgreiche Mikro-TENN-Entwicklung vorgestellten Dorsal Wurzelganglion (DRG) Neuronen 32 . Die Hydrogel-Mikrosäulen können erzeugt werden (Abbildung 2A ) durch Zugabe von flüssiger Agarose zu einer maßgeschneiderten, lasergeschnittenen zylindrischen Kanalanordnung oder zu Kapillarrohren, die beide ausgerichtete Akupunkturnadeln enthalten. Die Nadel bildet das Lumen und bestimmt den Innendurchmesser (ID) der Mikrosäule, während die Kapillarröhren-ID und der Durchmesser der Zylinder in der Laserschneidvorrichtung den Außendurchmesser (OD) der Konstrukte diktieren. Die OD und ID können entsprechend der gewünschten Anwendung gewählt werden, indem man unterschiedliche Durchmesser für die Geräte / Kapillarrohre bzw. die Akupunkturnadeln auswählt. Die Länge der Mikrosäulen kann auch variiert werden; Bisher haben wir den Bau von Mikro-TENNs bis zu 20 mm in der Länge 10 gemeldet und betreiben aktiv noch längere Längen. Nach den Agarosegelen und der Akupunktur nEedles werden entfernt, eine ECM-Lösung, die im allgemeinen aus Kollagen Typ I und Laminin besteht, wird dem Lumen der Konstrukte hinzugefügt ( Fig. 2C ). Der ECM-Kern sorgt für ein Gerüst zur Unterstützung der neuronalen Zelladhäsion und des axonalen Auswuchses. Anfänglich wurden primäre Ratten-kortikale Neuronen in den Mikrosäulen unter Verwendung von dissoziierten Zellsuspensionen 10 , 31 , 32 plattiert. Allerdings hat dieser Ansatz in allen Fällen nicht die Ziel-Cytoarchitektur hervorgebracht, die als die neuronalen Zellkörper definiert wurde, die auf die Enden der Mikrosäulen beschränkt waren, wobei das zentrale Lumen aus reinen, ausgerichteten axonalen Traktaten bestand. Seither hat die Verwendung eines erzwungenen neuronalen Aggregationsverfahrens (basierend auf Protokollen von Ungrin et al .) Eine zuverlässigere und konsistentere Herstellung von Mikro-TENNs mit der idealen Struktur ermöglicht (Abbildung 2B ) 44 . Neben der Beschreibung der aktuellenMethodik, wird dieser Artikel zeigen repräsentative Phasenkontrast und konfokale Bilder von Mikro-TENNs, die die Bildung von axonalen Traktaten im Laufe der Zeit, sowie die finalisierte Ziel-Zytoarchitektur zeigen. Dieses Manuskript wird auch auf bemerkenswerte Aspekte des Protokolls und die verbleibenden Herausforderungen und zukünftigen Richtungen der Mikro-TENN-Technologie erweitern.
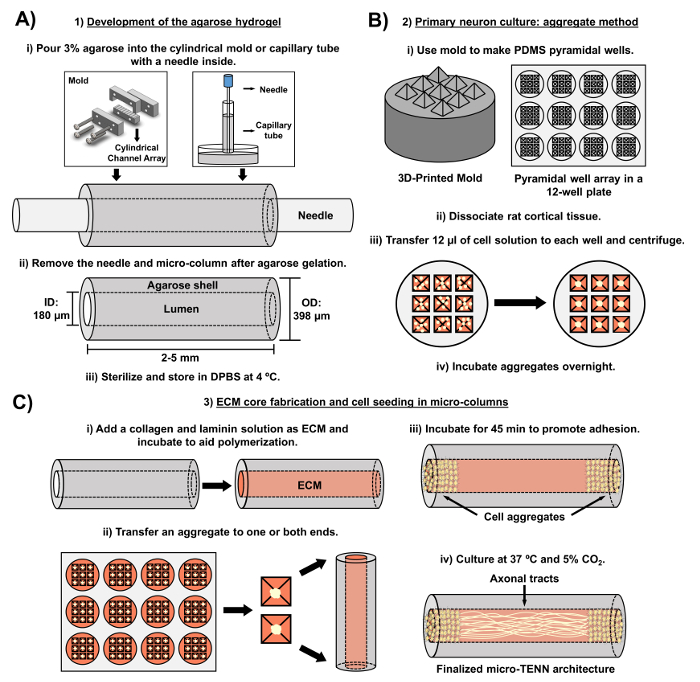
Abbildung 2: Schematische Darstellung des dreistufigen Mikro-TENN-Herstellungsprozesses. ( A ) Entwicklung des Agarose-Hydrogels: (i) Zunächst wird eine kleine Akupunkturnadel ( z. B. 180-350 μm Durchmesser) in die zylindrischen Kanäle einer maßgeschneiderten, lasergeschnittenen Form oder eines Kapillarrohres ( z , Durchmesser von 380-700 μm). Im nächsten Schritt wird flüssige Agarose in DPBS in die zylindrischen Kanäle oder Kapillarrohre eingeführt. (Ii) Nach den Agarosegelen wird die Nadel entfernt undDie Form wird zerlegt, um die hohlen Agarose-Mikrosäulen zu ergeben. (Iii) Diese Konstrukte werden dann sterilisiert und in DPBS gelagert. ( B ) Primäre Neuronenkultur und die Aggregatmethode: (i) Die neuronale Aggregation wird in pyramidenförmigen Mikro-Well-Arrays durchgeführt, die aus 3D-bedruckten Formen gegossen werden, die in die Vertiefungen einer 12-Well-Kulturplatte passen. (Ii) Mikro-TENNs umfassen primäre Rattenneuronen, die von fetalen Gehirnen von Embryon-Tag-18-Ratten dissoziiert sind. Nach der Gewebedissoziation mit Trypsin-EDTA und DNase I wird eine Zelllösung mit einer Dichte von 1,0-2,0 x 10 & sup6; Zelle / ml hergestellt. (Iii) 12 & mgr; l dieser Lösung werden in jede Vertiefung in dem pyramidenförmigen Mikro-Well-Array übertragen. Die Platte, die diese Mikro-Vertiefungen enthält, wird zentrifugiert, um Zellaggregate zu erzeugen. (Iv) Diese werden dann über Nacht vor dem Plattieren in den Mikrosäulen inkubiert. ( C ) ECM-Kernherstellung und Zellseeding: (i) Vor der Zellseedung wurde eine ECM-Lösung mit 1 mg / ml Typ I Kollagen und 1 mg / mlLaminin wird in das Innere der Mikro-TENNs überführt und polymerisiert. (Ii) Abhängig davon, ob unidirektionale oder bidirektionale Mikro-TENNs hergestellt werden, wird ein Aggregat an einem oder beiden Extremen der Mikrosäule platziert. (Iii) Nach einer Inkubationszeit zur Förderung der Adhäsion werden Mikro-TENNs in Petrischalen gezüchtet, die mit einem ergänzten embryonalen neuronalen Basalmedium überflutet sind. (Iv) Nach 3-5 Tagen in Kultur sollte die endgültige Mikro-TENN-Struktur Zelle Aggregate an den Extremen der Mikrosäule zeigen, wobei axonale Traktate ihre Länge überspannen. Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
