Una caratteristica comune dei disturbi e delle malattie del sistema nervoso centrale (CNS) come la lesione traumatica del cervello (TBI), lesioni del midollo spinale (SCI), ictus, malattia di Alzheimer e la malattia di Parkinson è la disconnessione dei percorsi assonali e delle cellule neuronali Perdita 1 , 2 , 3 , 4 , 5 , 6 . Ad esempio, quando un colpo ischemico non viene trattato, si stima che gli assoni siano persi a una velocità di 7 miglia di assoni al minuto 5 . Nel caso di TBI, che circa 1,7 milioni di persone sperimentano ogni anno negli Stati Uniti, la degenerazione assonale può continuare a verificarsi anni dopo il trauma, in quanto il pregiudizio iniziale precipita uno stato neurodegenerativo a lungo termine 4 . Aggravatando questi effetti deleteri, il CNS ha una capacità fortemente limitataCittà per la rigenerazione 1 , 7 , 8 , 9 . Dopo la lesione, si sviluppa un ambiente inibitorio caratterizzato da una mancanza di guida diretta a bersagli distanti, nella presenza di inibitori associati alla mielina che ostacolano la crescita del neurite e la formazione di una cicatrice gliale mediante astrociti reattivi 8 , 10 , 11 , 12 . La cicatrice gliale serve come barriera biochimica e fisica alla rigenerazione, con molecole come proteoglicani di condroitina solfato che ostruiscono la crescita dell'assone 8 , 11 . Inoltre, anche se le cellule staminali neurali sono state trovate nel CNS adulto, la produzione di nuovi neuroni è limitata, in quanto una prova consistente di neurogenesi è stata trovata solo nel bulbo olfattivo, l'ippocampoZona subgranulare, zona periventricolare e canale centrale del midollo spinale 13 , 14 . Questi ostacoli impediscono il recupero funzionale dei neuroni perduti e dell'architettura della materia bianca dopo lesioni o malattie, con conseguente effetto spesso alterato e prolungato di queste condizioni.
Nonostante la mancanza di capacità rigenerativa nel CNS adulte, è stato dimostrato che la rigenerazione assonale è possibile se vengono presentati adeguati suggerimenti ambientali ai neuroni ospiti 15 , 16 , 17 , 18 . I ricercatori hanno cercato di trasportare e manipolare fattori di crescita ( ad esempio, fattore di crescita del nervo, fattore di crescita epidermico, fattore di crescita dipendente dagli ialti e fattore neurotrofico 3) e altre molecole di guida per stimolare la plasticità e la rigenerazione dell'assone 14 ,/ Sup> 18 , 19 . Anche se questi studi hanno confermato che gli assoni adulti sono in grado di rispondere ai fattori di crescita, queste strategie sono limitate dalla bassa permeabilità della barriera emato-sangue e dalle specifiche sfumature spaziali e temporali necessarie per promuovere la rigenerazione 14 , 18 e 19 . Altri approcci hanno affidato l'iperattivazione dei fattori di trascrizione correlati alla rigenerazione nei neuroni del CNS. Ad esempio, la sovraespressione del fattore di trascrizione Stat3 stimola la rigenerazione assonale nel nervo ottico 20 . Tuttavia, sia la consegna della biomolecola che la sovraespressione dei fattori di trascrizione non riescono a sostituire le popolazioni neuronali perse. Le strategie cellulari sono state principalmente incentrate sul trapianto di cellule staminali neurali (NSC) nel CNS, sfruttando la loro capacità di sostituire i neuroni CNS, rilasciare fattori trofici,E sostenere i tentativi di neurogenesi che si verificano dopo lesioni 17 . Nonostante ciò, ci sono ancora pressanti sfide che ostacolano questo approccio, compresa la capacità ostacolata delle cellule neurali trapiantate per sopravvivere, integrarsi con l'ospite e rimanere spazialmente limitati all'area ferita 6 , 14 , 17 , 21 . Inoltre, la sola consegna cellulare non è in grado di reintegrare la citoarchitettura di percorsi axonal danneggiati o persi. Un approccio alternativo che affronta i problemi che si trovano ad affrontare le strategie di consegna di cellule e droga / chimica sta combinando questi approcci con l'uso dei biomateriali 14 , 22 , 23 . I biomateriali come gli idrogeli sono in grado di emulare le proprietà biochimiche e fisiche della matrice extracellulare (ECM), aiutando la distribuzione cellulare eD all'interno dell'area ferita e fornendo fattori di crescita e altre molecole bioattive con rilascio controllato 22 . Le caratteristiche interessanti di queste strategie basate su biomateriali hanno portato alla prova della rigenerazione assonica in vivo dopo il trapianto di scaffolds alla zona lesionata 24 , 25 , 26 , 27 , 28 , 29 , 30 . Tuttavia, le strategie biomateriali acellulari non sostituiscono le popolazioni neuronali perse; Quando vengono utilizzati come veicoli di erogazione per cellule precursori neuronali, gliali o neuronali, i biomateriali non sono in grado di ricostituire reti assonali a lunga distanza. La sfida di sviluppare un approccio che affronta sia la degenerazione del percorso assonale che la perdita neuronale associati a lesioni e malattie del CNS rimangono ancora <Sup class = "xref"> 31.
Il nostro gruppo di ricerca ha riportato in precedenza lo sviluppo di reti neurali (micro-TENN), progettate da micropagnetismi impiantabili, che sono un tipo di "scaffold vivente" costituito da corpi cellulari neuronali limitati a una o entrambe le estremità di una micro-colonna idrogel-ECM agarosio , Con tratti assonali allineati in tutto l'interno di questo incasso tridimensionale (3D) 1 , 10 , 31 , 32 . Una delle principali differenze tra questa tecnica e gli approcci precedenti è che la citoarchitettura delle micro-TENNs è stata completamente creata in vitro e viene poi trapiantato dopo 33 , 34 , 35 , 36 , 37 , 38 ,Sup class = "xref"> 39 , 40 , 41 . La fabbricazione in vitro offre un ampio controllo spaziale e temporale del fenotipo cellulare e dell'orientamento, delle proprietà meccaniche / fisiche, dei segnali biochimici e dei fattori esogeni, che beneficiano dell'integrazione di questi scaffolds con l'ospite dopo l'impianto 41 , 42 . Le micro-TENN sono ispirate anatomicamente perché emulano la neuroanatomia del cervello, mostrando tracce assonali simili a quelle che ponte le distinte aree funzionali del cervello ( Figura 1A ) 1 . Pertanto, questa strategia potrebbe essere in grado di sostituire fisicamente i tratti di tessuti bianchi persi e neuroni dopo l'impianto in una regione lesionata. Questa tecnica è ispirata anche da meccanismi di sviluppo in cui "scaffolds viventi naturali" formati da cellule gliali radiali e assoni pionieristici agiscono come guide di percorrenza per celluleMigrazione dalla zona subventricolare e outgrowth axon, rispettivamente 43 . Questi meccanismi sono ricapitolati nei tratti assonali allineati di micro-TENN, che possono presentare percorsi di vita per la migrazione delle cellule neurali e la rigenerazione assonale mediante una crescita assonale mediata dall'assone ( Figura 1C ) 43 . Inoltre, questa strategia sfrutta l'integrazione sinaptica tra i neuroni micro-TENN e il circuito nativo, formando nuovi relè che possono contribuire al recupero funzionale ( Figura 1B ) 43 . La capacità di formazione di sinapsi può anche concedere a questo approccio la capacità di modulare il CNS e di rispondere al tessuto dell'ospite in base ai feedback di rete. Ad esempio, i neuroni ottogeneticamente attivi nei montanti viventi possono essere stimolati per modulare i neuroni dell'ospite attraverso interazioni sinaptiche ( Figura 1D ).
Inoltre, il costruttore tubolare a base biomaterialeLa presenza di micro-TENN offre un ambiente adeguato per l'adesione, la crescita, l'estensione del neurite e la segnalazione delle cellule, mentre le dimensioni in miniatura dei costrutti potenzialmente consentono l'impianto minima invasivo e forniscono un microambiente parzialmente sequestrato per una graduale integrazione nel cervello. Infatti, le pubblicazioni recenti hanno dimostrato il potenziale delle micro-TENN per imitare i percorsi neurali dopo l'impianto nel cervello del topo. A seguito di microinjection stereotassiale, abbiamo precedentemente riportato prove di sopravvivenza neuronale micro-TENN, mantenimento dell'architettura del tratto assonale e estensione del neurite nella corteccia ospite per almeno 1 mese in vivo 10 , 31 . Inoltre, l'etichettatura con la sinapsina ha fornito prove istologiche dell'integrazione sinaptica con il tessuto nativo 10 , 31 . Nel complesso, le micro-TENN possono essere particolarmente adattate per ricostruire e modulare danniCNS sostituendo i neuroni perduti, integrando sinapticamente con il circuito ospite, ripristinando la citoarchitettura assonale persa e, in alcuni casi, fornendo assoni rigeneranti con gli opportuni segnali di percorrenza.
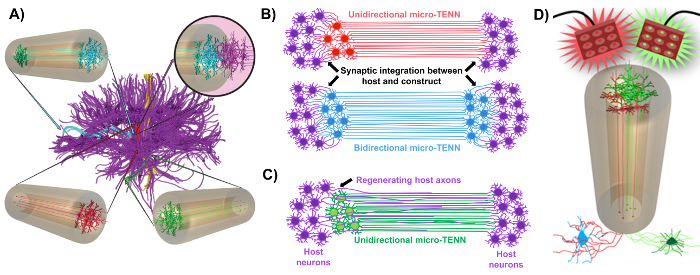
Figura 1: Principi e ispirazione dietro lo sviluppo di reti neurali innovative (micro-TENN). ( A ) Le micro-TENN simulano la citoarchitettura del connettore del cervello (viola), in cui regioni funzionalmente distinte sono collegate da tratti assonali lunghi e allineati in modo unidirezionale (rosso, verde) o bidirezionale (blu). Come esempio, le micro-TENN potrebbero ricostituire le connessioni perse in percorsi corticotalamici e nigrostriatali o nel percorso perforante dalla corteccia entorilina all'ippocampo (adattato da Struzyna et al. , 2015) 1 . ( B ) Diagramma di una unidirezioneL e bidimensionale micro-TENN (rosso e blu) sinapticamente integranti con il circuito ospite (viola) per servire come relè funzionale tra le due estremità di una lesione. ( C ) Schema dei tratti assonali di un micro-TENN unidirezionale (verde) che funge da guida per la rigenerazione degli assoni ospitanti (viola) facilitati dall'assone verso un obiettivo con cui il micro-TENN interagisce. ( D ) Schema concettuale dell'utilizzo di micro-TENNS attivo-otticamente attivi come neuromodulatori, sfruttando l'integrazione sinaptica con neuroni eccitatori o inibitori (in basso). Clicca qui per visualizzare una versione più grande di questa figura.
L'attuale manoscritto illustra la metodologia utilizzata per la realizzazione di micro-TENN utilizzando i neuroni corticali cerebrali embrionati. In particolare, le micro-TENN potrebbero essere fabbricate con altri tipi di cellule neurali. Per esAmpiamente, i primi rapporti di sviluppo micro-TENN di successo hanno caratterizzato i neuroni del ganglio radicale dorsale (DRG) 32 . Le micro-colonne idrogel possono essere generate ( Figura 2A ) aggiungendo agarosio liquido ad una matrice cilindrica a taglio laser su misura oa tubi capillari, entrambi contenenti aghi allineati di agopuntura. L'ago forma il lumen e determina il diametro interno (ID) della micro-colonna, mentre l'ID tubo capillare e il diametro dei cilindri nel dispositivo taglio laser dettano il diametro esterno (OD) dei costrutti. L'OD e l'ID possono essere scelti in base all'applicazione desiderata selezionando diametri differenti per i tubi del dispositivo / capillare e gli aghi di agopuntura rispettivamente. La lunghezza delle micro-colonne può anche essere variata; Ad oggi, abbiamo riportato la costruzione di micro-TENN fino a 20 mm di lunghezza 10 e perseguono attivamente lunghezze ancora più lunghe. Dopo i gel agarosici e l'agopuntura nLe eedle vengono rimosse, una soluzione ECM generalmente costituita da collagene di tipo I e laminina viene aggiunta al lumen dei costrutti ( figura 2C ). Il nucleo ECM fornisce un ponteggio per supportare l'adesione delle cellule neuronali e l'espansione assonica. Inizialmente i neuroni corticali primari del ratto venivano placcati nelle micro-colonne utilizzando sospensioni cellulari dissociate 10 , 31 , 32 . Tuttavia, questo approccio non ha prodotto la citoarchitettura target in tutti i casi, definita come i corpi delle cellule neuronali limitati alle estremità delle micro-colonne, con il lumen centrale costituito da tratti assonali puri allineati. Da allora, l'uso di un metodo di aggregazione neuronale forzata (basato su protocolli adattati da Ungrin e altri ) ha permesso una più affidabile e coerente fabbricazione di micro-TENNs con la struttura ideale ( figura 2B ) 44 . Oltre a descrivere la correnteMetodo, questo articolo mostrerà rappresentativi di fase-fase e immagini confocali di micro-TENN che dimostrano la formazione di tratti assonali nel tempo, così come la citoarchitettura finalizzata. Questo manoscritto si espanderà anche sugli aspetti degni di nota del protocollo e sulle sfide e sulle future direzioni della tecnologia micro-TENN.
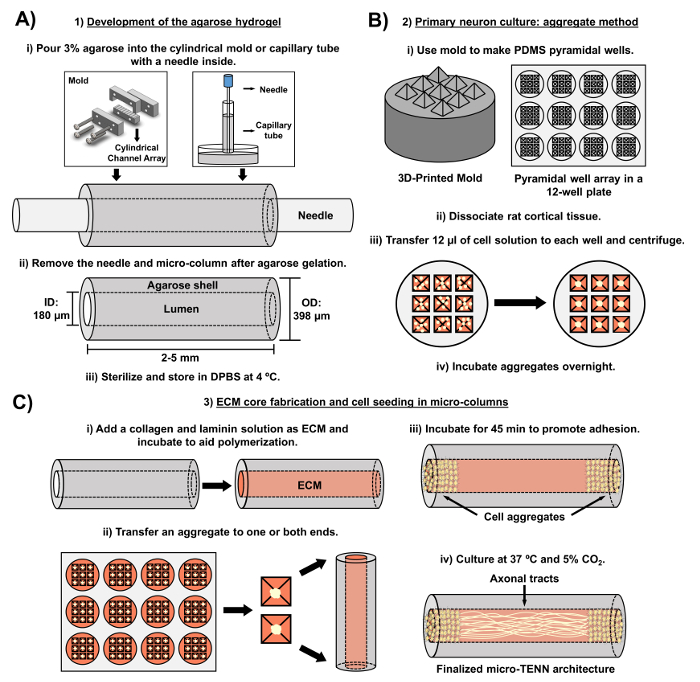
Figura 2: Diagramma schema del processo di fabbricazione micro-TENN a tre stadi. ( A ) Sviluppo dell'idrogel agarosico: (i) Inizialmente, un piccolo aghi di agopuntura ( ad esempio , diametro di 180-350 μm) viene inserito nei canali cilindrici di uno stampo a taglio laser su misura o di un capillare ( ad es. , Diametro 380-700 μm). Nella fase successiva, l'agarosio liquido in DPBS viene introdotto nei canali cilindrici o nei tubi capillari. (Ii) Dopo i gel di agarosio, l 'ago viene rimosso eLo stampo viene smontato per produrre le micro-colonne cave agarose. (Iii) Questi costrutti vengono poi sterilizzati e memorizzati in DPBS. ( B ) La cultura neuronale primaria e il metodo aggregato: (i) L'aggregazione neuronale viene eseguita in matrici di micro-pozzi piramidali, stampate da stampi stampati in 3D, che si inseriscono nei pozzetti di una piastra di coltura a 12 pozzetti. (Ii) Le micro-TENN includono i neuroni primari di ratto dissociati dai cervelli fetali dei ratti embrionali-giorno-18. Dopo la dissociazione tissutale con trypsina-EDTA e DNasi I, viene preparata una soluzione cellulare di densità 1,0-2,0 x 106 cellule / mL. Iii) 12 μL di questa soluzione vengono trasferiti a ciascun pozzetto nella matrice piramidale a micro-pozzetto. La piastra contenente questi micro-pozzetti viene centrifugata per produrre aggregati cellulari. (Iv) Questi vengono quindi incubati durante la notte prima della placcatura nelle micro-colonne. ( C ) Fabbricazione del nucleo ECM e sementi cellulari: (i) Prima della semina cellulare, una soluzione ECM contenente 1 mg / ml di collagene di tipo I e 1 mg / mlIl laminino viene trasferito all'interno delle micro-TENN e permesso di polimerizzare. (Ii) A seconda che si producano micro-TENN unidirezionali o bidirezionali, un aggregato viene posto rispettivamente in uno o entrambi gli estremi della micro-colonna. (Iii) Dopo un periodo di incubazione per promuovere l'adesione, le micro-TENN vengono coltivate in piatti Petri inondati con un mezzo basale neuronale embrionale integrato. (Iv) Dopo 3-5 giorni di coltura, la struttura finale del micro-TENN dovrebbe dimostrare gli aggregati di cellule agli estremi della micro-colonna, con tracce assonali che coprono la sua lunghezza. Clicca qui per visualizzare una versione più grande di questa figura.
