Общей характеристикой расстройств и заболеваний центральной нервной системы (ЦНС), таких как травматическая травма головного мозга (TBI), травма спинного мозга (SCI), инсульт, болезнь Альцгеймера и болезнь Паркинсона, является разъединение аксональных путей и нейронных клеток Потери 1 , 2 , 3 , 4 , 5 , 6 . Например, когда ишемический инсульт не лечится, считается, что аксоны теряются со скоростью 7 миль аксонов в минуту 5 . В случае TBI, который ежегодно испытывает приблизительно 1,7 миллиона человек в США, аксональная дегенерация может продолжаться и после нескольких лет после травмы, поскольку первоначальная травма ускоряет долгосрочное нейродегенеративное состояние 4 . Усугубляя эти вредные эффекты, ЦНС имеет строго ограниченный capaГород для регенерации 1 , 7 , 8 , 9 . После травмы развивается тормозная среда, которая характеризуется отсутствием направленного руководства к отдаленным мишеням, наличию ингибиторов, связанных с миелином, которые препятствуют росту нейритов и образованию глиального шрама реактивными астроцитами 8 , 10 , 11 , 12 . Глиальный шрам служит биохимическим и физическим барьером для регенерации, причем молекулы, такие как протеогликаны хондроитинсульфата, препятствуют росту аксонов 8 , 11 . Кроме того, несмотря на то, что нейронные стволовые клетки обнаружены во взрослой ЦНС, производство новых нейронов ограничено, поскольку последовательные доказательства нейрогенеза были обнаружены только в обонятельной луковице, гиппокампеСубгранулярная зона, перивентрикулярная область и центральный канал спинного мозга 13 , 14 . Эти препятствия препятствуют функциональному восстановлению потерянных нейронов и архитектуры белого вещества после травмы или заболевания, что приводит к часто меняющимся и продолжительным последствиям этих состояний.
Несмотря на отсутствие регенерационной способности у взрослых ЦНС, было продемонстрировано, что регенерация аксонов возможна, если адекватные экологические сигналы представлены в принимающих нейронах 15 , 16 , 17 , 18 . Исследователи пытались доставить и манипулировать факторами роста ( например, фактором роста нервов, эпидермальным фактором роста, глиально-зависимым фактором роста и нейротрофическим фактором-3) и другими молекулами наведения для стимулирования пластичности и регенерации аксонов 14 , </ Sup> 18 , 19 . Несмотря на то, что эти исследования подтвердили, что взрослые аксоны способны реагировать на факторы роста, эти стратегии ограничены низкой проницаемостью гематоэнцефалического барьера и специфическими пространственными и временными градиентами, необходимыми для стимулирования регенерации 14 , 18 , 19 . Другие подходы основывались на гиперактивации факторов транскрипции, связанных с регенерацией, в нейронах ЦНС. Например, сверхэкспрессия фактора транскрипции Stat3 стимулировала регенерацию аксонов в зрительном нерве 20 . Тем не менее, как доставка биомолекулы, так и избыточная экспрессия факторов транскрипции не позволяют заменить потерянные популяции нейронов. Клеточные стратегии в основном сосредоточены на трансплантации нервных стволовых клеток (НСК) в ЦНС, используя их способность заменять нейроны ЦНС, высвобождать трофические факторы,И поддерживать попытки нейрогенеза, которые возникают после травмы 17 . Несмотря на это, все еще существуют насущные проблемы, мешающие этому подходу, в том числе затрудненная способность пересаженных нейронных клеток выжить, интегрироваться с хозяином и оставаться пространственно ограничена поврежденной областью 6 , 14 , 17 , 21 . Кроме того, доставка клеток сама по себе неспособна восстановить цитоархитектуру поврежденных или потерянных аксональных путей. Альтернативный подход, который решает проблемы, стоящие перед стратегиями доставки клеток и наркотиков / химических веществ, сочетает эти подходы с использованием биоматериалов 14 , 22 , 23 . Биоматериалы, такие как гидрогели, способны эмулировать биохимические и физические свойства внеклеточного матрикса (ECM), помогая в доставке клетокD в пределах поврежденной области и доставки факторов роста и других биоактивных молекул с контролируемым высвобождением 22 . Привлекательные характеристики этих стратегий на основе биоматериалов привели к доказательству регенерации аксонов in vivo после трансплантации лесов в пораженные участки 24 , 25 , 26 , 27 , 28 , 29 , 30 . Однако стратегии бесклеточного биоматериала не заменяют потерянные нейронные популяции; При использовании в качестве средств доставки для нейронных, глиальных или нейронных клеток-предшественников, биоматериалы не способны восстанавливать сети аксонов на большие расстояния. Задача разработки подхода, который решает как дегенерацию аксонального пути, так и потерю нейронов, связанную с повреждением ЦНС и заболеванием, по-прежнему остается <Sup class = "xref"> 31.
Наша исследовательская группа ранее сообщала о разработке имплантируемых микро-тканевых нейронных сетей (micro-TENNs), которые являются типом «живого леса», состоящего из тел нейронных клеток, ограниченных одним или обоими концами микроколона с агарозным гидрогелем-ECM , С выровненными аксональными трактами, проходящими по всему внутреннему пространству этого трехмерного (3D) корпуса 1 , 10 , 31 , 32 . Одним из основных различий между этим методом и предыдущими подходами является то, что цитоархитектура микро-TENN создается полностью in vitro и затем трансплантируется 33 , 34 , 35 , 36 , 37 , 38 , <Sup class = "xref"> 39 , 40 , 41 . Изготовление in vitro предлагает обширный пространственный и временный контроль клеточного фенотипа и ориентации, механические / физические свойства, биохимические сигналы и экзогенные факторы, что выгодно для интеграции этих лесов с хозяином после имплантации 41 , 42 . Micro-TENN анатомически вдохновлены, потому что они эмулируют нейроанатомию мозга, отображая аксональные тракты, похожие на те, которые соединяют различные функциональные области мозга ( рис. 1A ) 1 . Таким образом, эта стратегия может быть в состоянии физически заменить потерянные участки белого вещества и нейроны после имплантации в поврежденную область. Этот метод также вдохновлен механизмами развития, в которых «естественные живые леса», образованные радиальными глиальными клетками и пионерскими аксонами, действуют в качестве направляющих пути для клетокМиграция из субвентрикулярной зоны и рост аксонов соответственно 43 . Эти механизмы рекапилируются в выровненных аксональных участках микро-TENN, которые могут представлять живые пути миграции нейронных клеток и регенерации аксонов с помощью аксоноподобного аксонового выроста ( рис. 1C ) 43 . Кроме того, в этой стратегии используется синаптическая интеграция между нейронами микро-TENN и собственной схемой, образуя новые реле, которые могут способствовать функциональному восстановлению ( рис. 1B ) 43 . Способность к образованию синапсов также может предоставить этому подходу способность модулировать ЦНС и реагировать на ткань хозяина в соответствии с сетевой обратной связью. Например, оптогенетически активные нейроны в живых каркасах могут стимулироваться для модуляции нейронных хозяев через синаптические взаимодействия ( рис. 1D ).
Кроме того, трубчатый контур на основе биоматериаловМикроэнтенозы обеспечивают адекватную среду для клеточной адгезии, роста, расширения нейритов и сигнализации, в то время как миниатюрные размеры конструкций потенциально допускают минимально инвазивную имплантацию и обеспечивают частично секвестрированную микроокружение для постепенной интеграции в мозг. Действительно, недавние публикации продемонстрировали потенциал микро-TENN для имитации нейронных путей после имплантации в мозг крысы. После стереотаксической микроинъекции мы ранее сообщали об обнаружении выживаемости нейронов микро-TENN, поддержании архитектуры аксонального тракта и расширении нейритов в корте головного мозга до, по меньшей мере, 1 месяца in vivo 10 , 31 . Более того, маркировка синапсином давала гистологические свидетельства синаптической интеграции с нативной тканью 10 , 31 . В целом, микро-TENN могут быть уникальными для восстановления и модуляции поврежденныхCNS путем замены потерянных нейронов, синаптической интеграции с хост-схемой, восстановления утраченной аксонной цитоархитектуры и, в некоторых случаях, обеспечения регенерации аксонов соответствующими сигналами пути.
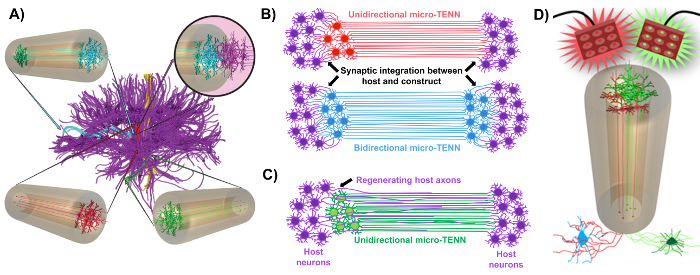
Рисунок 1: Принципы и вдохновение для разработки микросердечных нейронных сетей (микро-TENN). ( A ) Микро-TENN имитируют цитоархитектуру мозгового соединения (фиолетового), в котором функционально различные области связаны длинными выровненными аксональными трактами в однонаправленном (красном, зеленом) или двунаправленном (синем) порядке. Например, микро-TENN могут восстанавливать утраченные связи в кортикоталамическом и нигростриаторном путях или в перфорантном пути от энторинальной коры до гиппокампа (адаптировано из Struzyna et al. , 2015) 1 . ( B ) Диаграмма однонаправленногоL и двунаправленный микро-TENN (красный и синий соответственно), синаптически интегрируемые с главной схемой (фиолетовый), чтобы служить функциональным реле между обоими концами поражения. ( C ) Схема аксональных трактов однонаправленного микро-TENN (зеленая), служащая в качестве ориентира для облегченной аксоном регенерации аксонов хозяина (фиолетового) в направлении мишени, с которой взаимодействует микро-TENN. ( D ) Концептуальная схема использования оптико-активных микро-TENNS в качестве нейромодуляторов, с использованием синаптической интеграции с возбуждающими или тормозящими нейронами (снизу). Нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
В настоящей рукописи подробно изложена методология, используемая для изготовления микро-TENN с использованием эмбрионально полученных церебральных кортикальных нейронов. Примечательно, что микро-TENN могут быть изготовлены с использованием других типов нейронных клеток. ДляДостаточно, исходные сообщения об успешной разработке микро-TENN показали нейроны дорзального корня ганглия (DRG) 32 . Микроэлементы гидрогеля могут быть сгенерированы ( рис. 2А ) путем добавления жидкой агарозы к изготовленной на заказ цилиндрической канальной решетке с лазером или к капиллярным трубкам, содержащим выровненные иглы для иглоукалывания. Игла образует просвет и определяет внутренний диаметр (ID) микроколоны, в то время как идентификатор капиллярной трубки и диаметр цилиндров в устройстве для лазерной резки определяют внешний диаметр (OD) конструкций. OD и ID могут быть выбраны в соответствии с желаемым применением путем выбора различных диаметров для устройства / капиллярных труб и игл для иглоукалывания соответственно. Также можно варьировать длину микроколонок; На сегодняшний день мы сообщили о строительстве микро-ТЭНД длиной до 20 мм в длину 10 и активно преследуем еще более длинные длины. После агарозных гелей и иглоукалывания nЭдельы удаляются, к просвету конструкций добавляется раствор ECM, обычно состоящий из коллагена I типа и ламинина ( фиг. 2C ). Ядро ECM обеспечивает эшафот для поддержки адгезии нейронных клеток и роста аксонов. Первоначально первичные крысиные нейроны крыс были высеяны в микроколонах с использованием диссоциированных клеточных суспензий 10 , 31 , 32 . Однако этот подход не вызывал целевой цитоархитектуры во всех случаях, которая определялась как тела нейронных клеток, ограниченные концами микроколонок, причем центральный просвет состоял из чистых выровненных аксональных трактов. С тех пор использование метода агрегации принудительных нейронов (на основе протоколов, адаптированных из Ungrin и др .) Позволило более надежную и последовательную разработку микро-TENN с идеальной структурой ( рис. 2B ) 44 . В дополнение к описанию текущегоМетодологии, в этой статье будут представлены типичные фазоконтрастные и конфокальные изображения микро-TENN, которые демонстрируют формирование аксональных трактов с течением времени, а также завершенную целевую цитоархитектуру. Эта рукопись также расширит заслуживающие внимания аспекты протокола и остающиеся проблемы и будущие направления технологии микро-TENN.
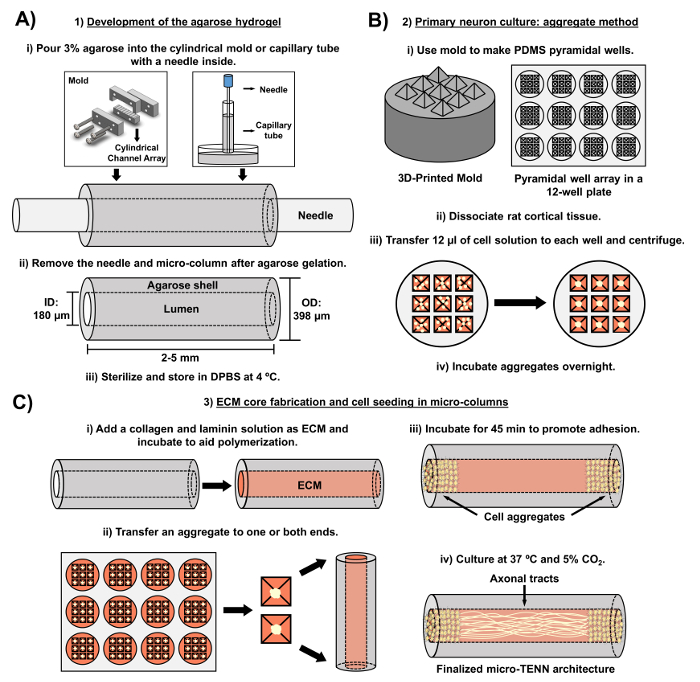
Рисунок 2: Принципиальная схема трехступенчатого процесса изготовления микро-TENN. ( A ) Разработка агарозного гидрогеля: (i) первоначально небольшая иглоукалывающая игла ( например , диаметр 180-350 мкм) вставляется в цилиндрические каналы изготовленной на заказ пресс-формы для лазерной резки или капиллярной трубки ( например, , 380-700 мкм в диаметре). На следующем этапе жидкая агароза в DPBS вводится в цилиндрические каналы или капиллярные трубки. (Ii) После агарозных гелей игла удаляется иПресс-форму разобрали, чтобы получить полые агарозные микроколоны. (Iii) Эти конструкции затем стерилизуют и хранят в DPBS. ( B ) Первичная культура нейронов и агрегированный метод: (i) Агрегирование нейронов выполняется в пирамидальных массивах микроячейки, отлитых из трехмерных печатных форм, которые помещаются в лунки 12-луночного планшета. (Ii) Микро-TENN включают первичные нейроны крыс, диссоциированные от эмбриональных мозгов крыс эмбрионального дня-18. После диссоциации ткани трипсином-EDTA и ДНКазой I готовят клеточный раствор с плотностью 1,0-2,0 × 10 6 клеток / мл. (Iii) 12 мкл этого раствора переносят в каждую лунку в пирамидальной микроячейке. Пластину, содержащую эти микро-лунки, центрифугируют для получения клеточных агрегатов. (Iv) Затем их инкубируют в течение ночи перед нанесением покрытия в микроколонах. ( C ) Изготовление сердечника ECM и клеточного посева: (i) Перед посевом клеток, ECM-раствор, содержащий 1 мг / мл коллагена I типа и 1 мг / млЛаминин переносят внутрь микро-TENN и дают возможность полимеризоваться. (Ii) В зависимости от того, изготавливаются ли однонаправленные или двунаправленные микро-TENN, агрегат помещается в одну или обе крайности микроколона соответственно. (Iii) После периода инкубации для содействия адгезии микро-TENN культивируют в чашках Петри, залитых добавленной эмбриональной нейронной базальной средой. (Iv) Через 3-5 дней в культуре окончательная микро-TENN-структура должна демонстрировать клеточные агрегаты в экстремумах микроколона, с аксональными трактами, охватывающими ее длину. Нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
