Een algemeen kenmerk van aandoeningen en ziekten van het centrale zenuwstelsel (CNS), zoals traumatisch hersenletsel (TBI), ruggenmergletsel (SCI), beroerte, de ziekte van Alzheimer en de ziekte van Parkinson, is de ontkoppeling van axonale wegen en neuronale cellen Verlies 1 , 2 , 3 , 4 , 5 , 6 . Bijvoorbeeld, wanneer een ischemische beroerte onbehandeld wordt, wordt geschat dat axonen verloren gaan met een snelheid van 7 mijl axonen per minuut 5 . In het geval van TBI, die ongeveer 1,7 miljoen mensen elk jaar alleen in de VS ervaren, kan axonale degeneratie jaar na trauma blijven optreden, aangezien de aanvankelijke verwonding een langdurige neurodegeneratieve toestand 4 neigt. Versterking van deze schadelijke effecten, het CNS heeft een zeer beperkte capaStad voor regeneratie 1 , 7 , 8 , 9 . Na aanleiding van het letsel ontstaat een remmende omgeving die wordt gekenmerkt door een gebrek aan gerichte begeleiding naar verre doelen, de aanwezigheid van myeline geassocieerde remmers die neuritale uitgroei verhinderen en de vorming van een glialeire door reactieve astrocyten 8 , 10 , 11 , 12 . Het gliaire doet dienst als een biochemische en fysieke barrière voor regeneratie, met moleculen zoals chondroïtinesulfaatproteoglycanen die axongroei 8 , 11 belemmeren. Bovendien, hoewel neurale stamcellen zijn gevonden in het volwassen CNS, is de productie van nieuwe neuronen beperkt, aangezien consistent bewijs van neurogenese alleen is gevonden in de olfactorische bol, de hippocampaleSubgranulaire zone, het periventriculaire gebied en het centrale kanaal van het ruggenmerg 13 , 14 . Deze obstakels verhinderen het functionele herstel van verloren neuronen en witte materie architectuur als gevolg van letsel of ziekte, wat resulteert in de vaak veranderende en langdurige effecten van deze omstandigheden.
Ondanks het gebrek aan regeneratieve capaciteit in het volwassen CNS, is aangetoond dat axonale regeneratie mogelijk is als adequate milieueisen worden voorgesteld aan de gastheer neuronen 15 , 16 , 17 , 18 . Onderzoekers hebben geprobeerd groeifactoren te leveren en te manipuleren ( bijv. De zenuwgroeifactor, de epidermale groeifactor, de glial-afhankelijke groeifactor en de neurotrofische factor-3) en andere begeleidingsmoleculen om de plasticiteit en axonregeneratie te stimuleren 14 , </ Sup> 18 , 19 . Hoewel deze studies hebben bevestigd dat volwassen axonen in staat zijn om op groeifactoren te reageren, worden deze strategieën beperkt door de lage permeabiliteit van de bloed-hersenbarrière en de specifieke ruimtelijke en temporale gradiënten die nodig zijn om regeneratie 14 , 18 , 19 te bevorderen. Andere benaderingen zijn gebaseerd op de hyperactivatie van regeneratie gerelateerde transcriptiefactoren in CNS neuronen. Bijvoorbeeld stimuleerde overexpressie van de stat3 transcriptiefactor axonale regeneratie in de optische zenuw 20 . Desalniettemin slaan zowel de biomoleculeafgifte als de overexpressie van transcriptiefactoren in de plaats van verloren neuronale populaties. Cell-based strategieën hebben vooral betrekking op transplantatie van neurale stamcellen (NSC's) in het CNS, waarbij zij profiteren van hun capaciteit om CNS neuronen te vervangen, trofische factoren te vrijgeven,En ondersteun de pogingen op neurogenese die zich voordoen na verwonding 17 . Ondanks dit zijn er nog steeds uitdagende uitdagingen die deze aanpak verhinderen, inclusief het belemmerde vermogen van getransplanteerde neurale cellen om te overleven, integreren met de gastheer en blijven ruimtelijk beperkt tot het gewond gebied 6 , 14 , 17 , 21 . Daarnaast is de levering van cellen alleen niet in staat om de cytoarchitectuur van beschadigde of verloren axonale wegen te herstellen. Een alternatieve benadering die betrekking heeft op de problemen die cel- en drug / chemische afleverstrategieën voordoen, combineert deze benaderingen met het gebruik van biomaterialen 14 , 22 , 23 . Biomaterialen zoals hydrogelen kunnen de biochemische en fysische eigenschappen van de extracellulaire matrix (ECM) emulerenD retentie binnen het gewond gebied, en het leveren van groeifactoren en andere bioactieve moleculen met gereguleerde afgifte 22 . De aantrekkelijke kenmerken van deze biomaterialen gebaseerde strategieën hebben geleid tot bewijs van in vivo axonale regeneratie na de transplantatie van steigers naar het lesion gebied 24 , 25 , 26 , 27 , 28 , 29 , 30 . Echter, acellulaire biomaterialestrategieën vervangen geen verloren neuronale populaties; Bij gebruik als leveringsvoertuigen voor neuronale, gliale of neuronale precursorcellen, zijn biomaterialen niet in staat om axonale netwerken op lange afstand te reconstrueren. De uitdaging om een aanpak te ontwikkelen die zowel de axonale wegdegeneratie als neuronale verlies in verband met CNS-letsel en ziekte bestrijkt, blijft <Sup class = "xref"> 31.
Onze onderzoeksgroep heeft eerder de ontwikkeling van implanteerbare microweefselgemanipuleerde neurale netwerken (micro-TENN's) gemeld, die een type "levende steiger" zijn die bestaat uit neuronale cellichamen die beperkt zijn tot één of beide uiteinden van een agarose hydrogel-ECM microkolom , Met uitgelijnd axonale tracten die zich uitstrekken over het binnenste van deze driedimensionale (3D) omhulling 1 , 10 , 31 , 32 . Een van de belangrijkste verschillen tussen deze techniek en eerdere benaderingen is dat de cytoarchitectuur van micro-TENNEN volledig in vitro wordt gecreëerd en daarna 33 , 34 , 35 , 36 , 37 , 38 wordt getransplanteerd <Sup class = "xref"> 39 , 40 , 41 . In vitro fabricage biedt uitgebreide ruimtelijke en temporale controle van cellulaire fenotype en oriëntatie, mechanische / fysische eigenschappen, biochemische signalen en exogene factoren, die de integratie van deze steigers bij de gastheer na de implantatie 41 , 42 ten goede komen . Micro-Tenns zijn anatomisch geïnspireerd omdat ze neuroanatomie van de hersenen emuleren, waarbij axonale tracten worden weergegeven die vergelijkbaar zijn met die welke verschillende functionele gebieden van de hersenen gebruiken ( Figuur 1A ) 1 . Daarom kan deze strategie fysieke vervanging van verloren witte materiaalkanalen en neuronen na implantatie in een gewaarschelde regio vervangen. Deze techniek wordt ook geïnspireerd door ontwikkelingsmechanismen waarin "natuurlijke levende stellingen" gevormd door radiale gliale cellen en pioniers axonen fungeren als pathfinding gidsen voor celMigratie uit de subventriculaire zone en axonale uitgroei, respectievelijk 43 . Deze mechanismen worden teruggevonden in de uitgelijste axonale tracten van micro-TENNs, die leefwegen voor neurale celmigratie en axonale regeneratie kunnen voorstellen door axon-gemedieerde axonale uitgroei ( Figuur 1C ) 43 . Bovendien maakt deze strategie gebruik van synaptische integratie tussen de micro-TENN-neuronen en de inheemse schakelingen, waarbij nieuwe relais worden gevormd die kunnen bijdragen tot functioneel herstel ( Figuur 1B ) 43 . De capaciteit voor synapsvorming kan deze aanpak ook de mogelijkheid bieden om het CNS te moduleren en te reageren op gastheerweefsel volgens netwerk feedback. Optogenetisch actieve neuronen in de levende stellingen kunnen bijvoorbeeld gestimuleerd worden om gastheerneuronen te moduleren door middel van synaptische interacties ( Figuur 1D ).
Daarnaast heeft de biomaterial-gebaseerde buisvormige constrUitvoering van micro-TENNs biedt een adequate omgeving voor celadhesie, groei, neurietuitbreiding en signalering, terwijl de miniatuurafmetingen van de constructen mogelijk minimaal invasieve implantatie toestaan en een gedeeltelijk gesekwestreerde microomgeving voor geleidelijke integratie in de hersenen mogelijk maken. Inderdaad hebben recente publicaties het potentieel van micro-TENNEN aangetoond om neurale trajecten na implantatie in de ratbrein te nabootsen. Na stereotaxische microinjectie hebben we eerder bewijs van micro-TENN neuronale overleving, onderhoud van axonale tractarchitectuur en neurietuitbreiding in de gastheercortex tot tenminste 1 maand in vivo 10 , 31 gemeld. Bovendien gaf etikettering met synapsine histologisch bewijs van synaptische integratie met natief weefsel 10 , 31 . In het algemeen kunnen micro-TENNES uniek geschikt zijn om beschadigd te reconstrueren en te modulerenCNS door verloren neuronen te vervangen, synaptisch te integreren met gastheercircuits, herstel van verloren axonale cytoarchitectuur en, in bepaalde gevallen, het regenereren van axonen met de juiste pathfinding cues.
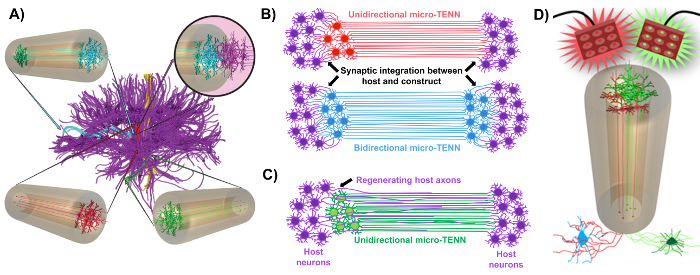
Figuur 1: Principes en inspiratie achter de ontwikkeling van micro-tissue engineered neurale netwerken (micro-TENNs). ( A ) Micro-Tenns nabootsen de cytoarchitectuur van de hersenverbindingen (paars), waarbij functionele afzonderlijke gebieden door middel van lange, uitgelijste axonale tracten in eenrichtingsbewerking (rood, groen) of tweerichtings (blauw) verbonden zijn. Bijvoorbeeld, micro-TENN's kunnen verloren verbindingen in corticothalamus- en nigrostriatale trajecten of in de perforante weg van de entorhinaire cortex tot de hippocampus reconstrueren (aangepast uit Struzyna et al. , 2015) 1 . ( B ) Diagram van een unidirectionaL en bidirectionele micro-TENN (respectievelijk rood en blauw) synaptisch integreren met de gastheercircuit (paars) om als een functioneel relais tussen beide uiteinden van een letsel te dienen. ( C ) Schematisch van de axonale tracten van een unidirectionele micro-TENN (groen), die dienen als een geleiding voor axon-gefaciliteerde regeneratie van host axons (paars) naar een doel waarmee de micro-TENN in wisselwerking is. ( D ) Conceptueel diagram van het gebruik van optogenetisch actieve micro-TENNS als neuromodulators, die profiteren van synaptische integratie met excitatoire of remmende neuronen (bodem). Klik hier om een grotere versie van deze figuur te bekijken.
Het huidige manuscript detaileert de methodologie die wordt gebruikt om micro-TENNEN te vervaardigen met behulp van embryonaal afgeleide cerebrale corticale neuronen. Met name kunnen micro-TENNEN worden vervaardigd met andere soorten neurale cellen. Voor exRuim, de initiële rapporten van succesvolle micro-TENN ontwikkeling gekenmerkt door dorsale root ganglion (DRG) neuronen 32 . De hydrogelmicrokolommen kunnen gegenereerd worden ( Figuur 2A ) door vloeibare agarose aan te voegen aan een op maat gemaakte, lasersnijdende cilindrische kanaalschikking of naar capillaire buizen, die beide uitgelijnd acupunctuurnaalden bevatten. De naald vormt het lumen en bepaalt de binnendiameter (ID) van de microkolom, terwijl de capillaire buis ID en de diameter van de cilinders in het lasergesneden apparaat de buitendiameter (OD) van de constructies dicteren. De OD en ID kunnen volgens de gewenste toepassing worden gekozen door respectievelijk verschillende diameters voor de inrichting / capillaire buizen en de acupunctuurnaalden te selecteren. De lengte van de microkolommen kan ook worden gevarieerd; Tot op heden hebben we de constructie van micro-TENNEN tot 20 mm lang 10 gerapporteerd en actief nog langere lengtes volgen. Na de agarose gels en de acupunctuur nEedles worden verwijderd, een ECM oplossing die in het algemeen bestaat uit type I collageen en laminine wordt toegevoegd aan het lumen van de constructen ( Figuur 2C ). De ECM-kern biedt een steiger ter ondersteuning van neuronale celadhesie en axonale uitgroei. Aanvankelijk werden primaire ratcorticale neuronen geplateerd in de microkolommen met behulp van gedissocieerde cel suspensies 10 , 31 , 32 . Deze aanpak produceerde echter niet in alle gevallen de target cytoarchitectuur, die werd gedefinieerd als de neuronale cellichamen die beperkt waren tot de uiteinden van de microkolommen, waarbij het centrale lumen bestaat uit zuivere uitgelijnd axonale tracten. Sindsdien is het gebruik van een gedwongen neuronale aggregatie methode (gebaseerd op protocols aangepast aan Ungrin et al .) Een betrouwbare en consistente fabricage van micro-TENNEN mogelijk gemaakt met de ideale structuur ( Figuur 2B ) 44 . Naast het beschrijven van de huidigeMethodologie, zal dit artikel representatieve fasecontrast- en confocale beelden van micro-TENNs tonen die de vorming van axonale tracten over de tijd tonen, evenals de gefinaliseerde doelcytoorarchitectuur. Dit manuscript zal ook uitbreiden op opmerkelijke aspecten van het protocol en de resterende uitdagingen en toekomstige aanwijzingen van de micro-TENN-technologie.
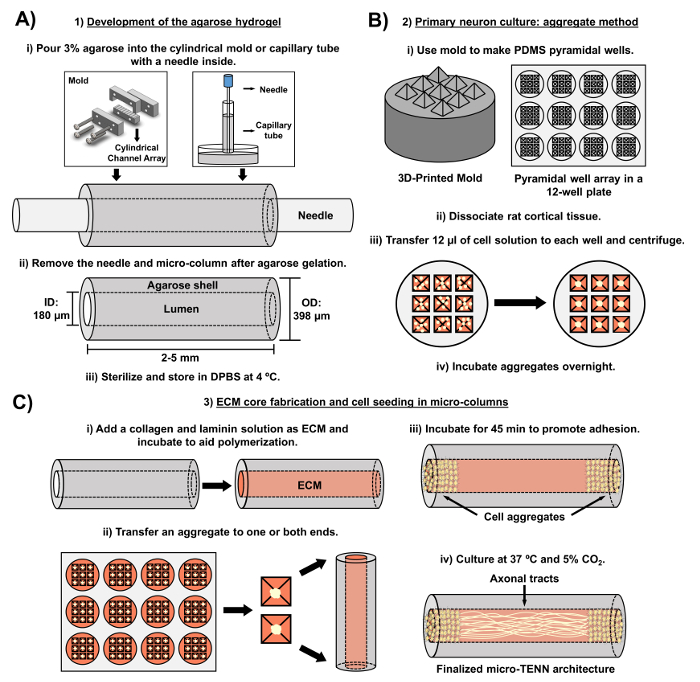
Figuur 2: Schematisch diagram van het driedimensionale micro-TENN fabricageproces. ( A ) Ontwikkeling van de agarosehydrogel: (i) Aanvankelijk wordt een kleine acupunctuurnaald ( bijv . 180-350 μm in diameter) in de cilindrische kanalen van een op maat gemaakte, lasergesneden vorm of een kapillair buis geplaatst ( bijv. , Diameter 380-700 μm). In de volgende stap wordt vloeibare agarose in DPBS geïntroduceerd in de cilindrische kanalen of capillaire buizen. (Ii) Na de agarosegels wordt de naald verwijderd enDe vorm wordt gedemonteerd om de holle agarose microkolommen te geven. (Iii) Deze constructies worden dan gesteriliseerd en opgeslagen in DPBS. ( B ) Primaire neuroncultuur en de aggregaatmethode: (i) Neuronale aggregatie wordt uitgevoerd in pyramidale micro-well arrays, gegoten uit 3D-gedrukte schimmels, die passen in de putjes van een 12-wells kweekplaat. (Ii) Micro-TENNs omvatten primaire ratneuronen die zijn losgelaten van foetale hersenen van embryonale-dag-18 ratten. Na afloop van weefseldissociatie met trypsine-EDTA en DNase I wordt een celoplossing met een dichtheid van 1,0-2,0 x 106 cellen / ml bereid. (Iii) 12 μl van deze oplossing worden overgebracht naar elke put in de pyramidale micro-put-array. De plaat die deze microputjes bevat, wordt gecentrifugeerd om celaggregaten te produceren. (Iv) Deze worden vervolgens overnacht geïncubeerd voorafgaand aan plating in de microkolommen. ( C ) ECM-kernfabrikatie en celzaden: (i) Vóór het zaaien van een cel, een ECM-oplossing die 1 mg / ml type I collageen en 1 mg / ml bevatLaminine wordt overgebracht naar het interieur van de micro-TENNEN en mag polymeriseren. (Ii) Afhankelijk van de vraag of unidirectionele of bidirectionele micro-TENNEN worden vervaardigd, wordt een aggregaat geplaatst op respectievelijk één of beide uitersten van de microkolom. Iii) Na een periode van incubatie om adhesie te bevorderen, worden micro-TENNEN gekweekt in Petri-schalen die zijn overspoeld met aangevulde embryonale neuronale basale media. Iv) Na 3-5 dagen in de cultuur, moet de uiteindelijke micro-TENN-structuur celaggregaten aantonen bij de uitersten van de microkolom, met axonale tracten die zijn lengte overspannen. Klik hier om een grotere versie van deze figuur te bekijken.
