En vanlig egenskap for lidelser og sykdommer i sentralnervesystemet (CNS), som traumatisk hjerneskade (TBI), ryggmargsskade (SCI), hjerneslag, Alzheimers sykdom og Parkinsons sykdom, er frakoblingen av aksonale veier og nevroncelle Tap 1 , 2 , 3 , 4 , 5 , 6 . For eksempel, når et iskemisk slag går ubehandlet, anslås det at aksonene går tapt med en hastighet på 7 miles av axoner per minutt 5 . I tilfelle av TBI, som ca 1,7 millioner mennesker opplever hvert år i USA alene, kan aksonal degenerasjon fortsette å skje år etter traumer, da den første skaden utsettes for en langsiktig neurodegenerativ tilstand 4 . Forverrer disse skadelige effektene, har CNS en alvorlig begrenset capaBy for regenerering 1 , 7 , 8 , 9 . Etter skader utvikles et hemmende miljø som er preget av mangel på rettet veiledning til fjerne mål, tilstedeværelsen av myelin-assosierte hemmere som hindrer nevittutvæksten, og dannelsen av et glialær ved reaktive astrocytter 8 , 10 , 11 , 12 . Glialæret tjener som en biokjemisk og fysisk barriere for regenerering, med molekyler som kondroitinsulfatproteoglykaner som hindrer aksonutvokst 8 , 11 . Videre, selv om neurale stamceller er blitt funnet i den voksne CNS, er produksjonen av nye nevroner begrenset, da konsekvent bevis på nevrogenese kun er funnet i olfaktorisk pære, hippocampalSubgranulær sone, det periventrikulære området og den sentrale kanal i ryggmargen 13 , 14 . Disse hindringene hindrer den funksjonelle gjenoppretting av tapt nevroner og hvit materiell arkitektur etter skade eller sykdom, noe som resulterer i de ofte endringsmessige og langvarige effektene av disse forholdene.
Til tross for mangel på regenerativ kapasitet i den voksne CNS, har det vist seg at aksonal regenerering er mulig dersom tilstrekkelige miljøveiledninger presenteres for verten nevroner 15 , 16 , 17 , 18 . Forskere har forsøkt å levere og manipulere vekstfaktorer (for eksempel nervevækstfaktor, epidermal vekstfaktor, glialavhengig vekstfaktor og nevrotrofisk faktor 3) og andre veiledningsmolekyler for å stimulere plastisitet og axonregenerering 14 , </ Sup> 18 , 19 . Selv om disse studiene har bekreftet at voksenaksoner er i stand til å reagere på vekstfaktorer, er disse strategiene begrenset av lavt gjennomtrengelighet av blod-hjernebarrieren og de spesifikke romlige og tidsmessige gradienter som kreves for å fremme regenerering 14 , 18 , 19 . Andre tilnærminger har stått på hyperaktivering av regenereringsrelaterte transkripsjonsfaktorer i CNS-neuroner. For eksempel stimulerte overekspresjon av Stat3 transkripsjonsfaktoren aksonal regenerering i optisk nerve 20 . Likevel unnlater både biomolekylavgivelse og overekspresjon av transkripsjonsfaktorer å erstatte tapte nevronpopulasjoner. Cell-baserte strategier har hovedsakelig sentrert seg på transplantasjon av neuralstamceller (NSC) i CNS, og utnytter deres evne til å erstatte CNS-neuroner, frigjøre trofiske faktorer,Og støtte forsøk på neurogenese som oppstår etter skade 17 . Til tross for dette er det fortsatt presserende utfordringer som hindrer denne tilnærmingen, inkludert den hindrede evne til transplanterte nevrale celler til å overleve, integrere med verten, og forblir romlig begrenset til det skadede område 6 , 14 , 17 , 21 . I tillegg er cellelevering alene ikke i stand til å gjenopprette cytoarkitekturen av skadede eller tapte aksonale veier. En alternativ tilnærming som løser problemene med celle- og stoff- / kjemisk leveringsstrategier, er å kombinere disse tilnærmingene med bruken av biomaterialene 14 , 22 , 23 . Biomaterialer som hydrogeler er i stand til å emulere de biokjemiske og fysiske egenskapene til den ekstracellulære matriksen (ECM), som bidrar til cellelevering ogD oppbevaring i det skadede området, og levere vekstfaktorer og andre bioaktive molekyler med kontrollert frigivelse 22 . De attraktive egenskapene til disse biomaterialebaserte strategiene har resultert i bevis på in vivo axonal regenerering etter transplantasjon av stillaser til det lesioned området 24 , 25 , 26 , 27 , 28 , 29 , 30 . Imidlertid erstatter acellulære biomaterialestrategier ikke tapte nevronpopulasjoner; Når de brukes som leveringsvogner for nevron-, glial- eller neuronprekursorceller, er biomaterialer ikke i stand til å rekonstruere langdistansaksonale nettverk. Utfordringen med å utvikle en tilnærming som takler både den aksonale banedegenerasjonen og nevrolog tap forbundet med CNS-skade og sykdom forblir fortsatt <Sup class = "xref"> 31.
Vår forskningsgruppe rapporterte tidligere utviklingen av implanterbare mikro-vev-konstruerte nevrale nettverk (mikro-tenner), som er en type "levende stillas" bestående av neuronelle celleorganer begrenset til en eller begge ender av en agarosehydrogel-ECM mikrosøjle , Med justerte aksonale kanaler som strekker seg gjennom det indre av denne tredimensjonale (3D) innkapslingen 1 , 10 , 31 , 32 . En av de viktigste forskjellene mellom denne teknikken og tidligere tilnærminger er at cytoarkitektur av mikro-tenner er opprettet helt in vitro og transplanteres etterpå 33 , 34 , 35 , 36 , 37 , 38 , <Sup class = "xref"> 39 , 40 , 41 . In vitro- fremstilling gir omfattende romlig og tidsmessig kontroll av cellulær fenotype og orientering, mekaniske / fysiske egenskaper, biokjemiske tegn og eksogene faktorer, noe som fordeler integrasjonen av disse stillasene med verten etter implantasjon 41 , 42 . Mikro-tenner er anatomisk inspirert fordi de etterligner hjernens neuroanatomi, viser aksonale kanaler som ligner de som knytter distinkte funksjonsområder av hjernen ( figur 1A ) 1 . Derfor kan denne strategien fysisk erstatte tapte hvite materielle kanaler og nevroner etter implantasjon i en lesioned region. Denne teknikken er også inspirert av utviklingsmekanismer der "naturlige levende stillaser" dannet av radiale glialceller og banebrytende axoner fungerer som veiledende guider for celleMigrasjon fra subventricular sonen og aksonal utvekst, henholdsvis 43 . Disse mekanismene er rekapitulert i de justerte aksonale kanaler av mikro-tenner, som kan presentere levende veier for nevralcellemigrasjon og aksonal regenerering ved akson-mediert aksonal utvekst ( figur 1C ) 43 . Videre utnytter denne strategien seg av synaptisk integrasjon mellom mikro-TENN-neuronene og det nasjonale kretsløpet, og danner nye reléer som kan bidra til funksjonell gjenoppretting ( figur 1B ) 43 . Kapasiteten til synapsdannelse kan også gi denne tilnærmingen muligheten til å modulere CNS og reagere på vertsvev i henhold til nettverks tilbakemelding. For eksempel kan optogenetisk aktive nevroner i levende stillasene stimuleres til å modulere vertsneuroner gjennom synaptiske interaksjoner ( Figur 1D ).
I tillegg er den biomaterialebaserte rørformede konstrUgn av mikro-tenner gir et tilstrekkelig miljø for celleadhesjon, vekst, neurittforlengelse og signalering, mens de minste dimensjonene av konstruksjonene muligens muliggjør minimal invasiv implantering og gir et delvis sekvestrert mikromiljø for gradvis integrering i hjernen. Faktisk har nyere publikasjoner vist potensialet for mikro-tenner til å etterligne nevrale veier etter implantasjon i rottehjerne. Etter stereotaksisk mikroinjeksjon rapporterte vi tidligere bevis for mikro-TENN-neuronal overlevelse, vedlikehold av aksonalkanalarkitektur, og nevittforlengelse i vertscortexen i minst 1 måned in vivo 10 , 31 . Dessuten ga merking med synapsin histologisk bevis på synaptisk integrasjon med nativt vev 10 , 31 . Samlet sett kan mikro-tenner være unikt egnet til å rekonstruere og modulere skadetCNS ved å erstatte tapte nevroner, synaptisk integrering med vertskretsløp, gjenopprette tapt axonal cytoarkitektur og i visse tilfeller å gi regenererende axoner med de riktige stikkordene.
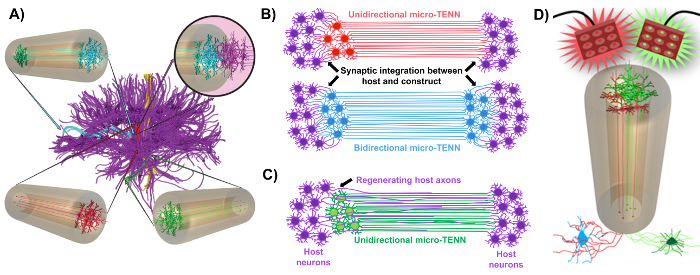
Figur 1: Prinsipper og inspirasjon bak utviklingen av mikro-vev-konstruerte nevrale nettverk (mikro-tenner). ( A ) Mikro-Tenner etterligner cytoarkitekturen til hjernekoblingen (lilla), hvor funksjonelt forskjellige regioner er forbundet med lange, justerte aksonale kanaler i en retningsriktet (rød, grønn) eller toveis (blå) måte. Som et eksempel kan mikro-tenner rekonstruere tapte forbindelser i kortikothalamiske og nigrostriatale veier eller i perforantveien fra entorhinal cortex til hippocampus (tilpasset fra Struzyna et al. , 2015) 1 . ( B ) Diagram over en enhetlig retningL og toveis mikro-TENN (henholdsvis rød og blå) synaptisk integrering med vertskretsen (lilla) for å fungere som et funksjonsrelé mellom begge ender av en lesjon. ( C ) Skjematisk av axonaltraktene i en ensrettet mikro-TENN (grønn) som tjener som en veiledning for aksonforenklet regenerering av vertsaxoner (lilla) mot et mål som mikro-tennene samhandler med. ( D ) Konseptbasert diagram for bruk av optogenetisk aktive mikro-TENNS som nervemodulatorer, utnytte synaptisk integrasjon med eksitatoriske eller hemmerende nevroner (bunn). Vennligst klikk her for å se en større versjon av denne figuren.
Det nåværende manuskriptet beskriver metoden som ble benyttet for å fremstille mikro-tenner ved bruk av embryonalt avledede cerebrale kortikale nevroner. Spesielt kan mikro-tenner fremstilles med andre typer nevrale celler. For eksempelRikelig, de første rapportene om vellykket mikro-TENN-utvikling inneholdt dorsal root ganglion (DRG) nevroner 32 . Hydrogelmikrokolonnene kan genereres ( figur 2A ) ved å tilsette flytende agarose til en skreddersydd, laser-kuttet sylindrisk kanalgruppe eller til kapillærrør, begge inneholdende justerte akupunkturnåler. Nålen danner lumenet og bestemmer mikrokolonnens indre diameter (ID), mens kapillærrøret ID og diameteren av sylindrene i den laserskårne anordningen dikterer den ytre diameter (OD) til konstruksjonene. OD og ID kan velges i henhold til ønsket søknad ved å velge forskjellige diametre for henholdsvis anordning / kapillærrør og akupunktur nåler. Lengden på mikrokolonnene kan også varieres; Til dags dato har vi rapportert bygging av mikro-tenner opptil 20 mm i lengde 10 og driver aktivt lengre lengder. Etter agarose geler og akupunktur nEedler fjernes, en ECM-løsning som vanligvis består av type I kollagen og laminin tilsettes i konstruksjonslumenet ( figur 2C ). ECM-kjernen gir et stillas for å understøtte nevroncelleadhesjon og aksonal utvekst. I første omgang ble primære rotte-kortikale nevroner plettet i mikrokolonnene ved anvendelse av dissocierte cellesuspensjoner 10 , 31 , 32 . Imidlertid produserte denne tilnærmingen ikke mål-cytoarkitekturen i alle tilfeller, som ble definert som neuronelle celleorganer begrenset til endene av mikrokolonnene, med det sentrale lumen bestående av renjusterte aksonale kanaler. Siden da har bruken av en tvungen neuronal aggregeringsmetode (basert på protokoller tilpasset Ungrin et al .) Gjort det mulig for en mer pålitelig og konsekvent fremstilling av mikro-tenner med den ideelle strukturen ( figur 2B ) 44 . I tillegg til å beskrive dagensMetodikk, vil denne artikkelen vise representative fasekontrast og konfokale bilder av mikro-tenner som demonstrerer dannelsen av aksonale kanaler over tid, samt den ferdige mål-cytoarkitekturen. Dette manuskriptet vil også utvide på bemerkelsesverdige aspekter av protokollen og gjenværende utfordringer og fremtidige retninger av mikro-TENN teknologien.
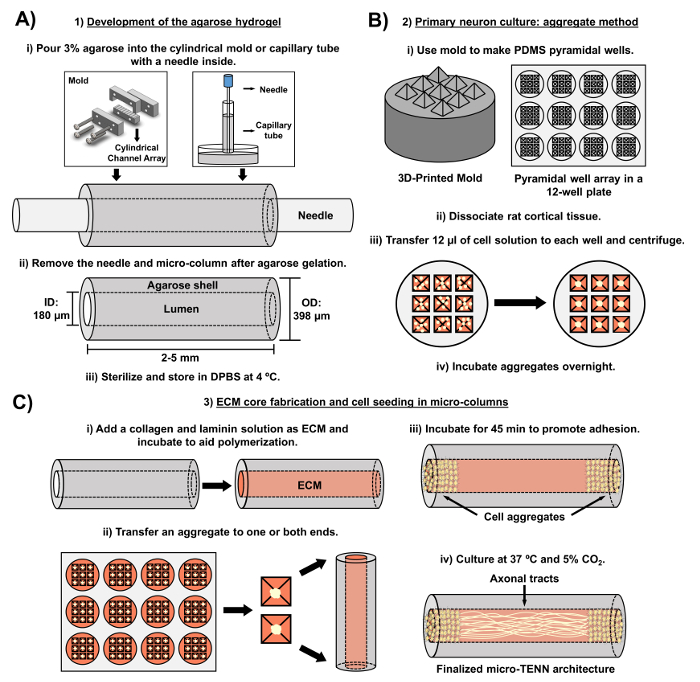
Figur 2: Skjematisk diagram av tre-trinns mikro-TENN fabrikasjonsprosess. ( A ) Utvikling av agarosehydrogelen: (i) Først settes en liten akupunkturnål ( f.eks . 180-350 μm i diameter) inn i de sylindriske kanalene i en skreddersydd, laserskåret form eller et kapillarrør ( f.eks. , Diameter 380-700 μm). I det neste trinnet blir flytende agarose i DPBS introdusert i de sylindriske kanalene eller kapillærrørene. (Ii) Etter agarose gelene fjernes nålen ogFormen er demontert for å gi de hule agarosemikolonner. (Iii) Disse konstruksjonene steriliseres deretter og lagres i DPBS. ( B ) Primærnekonkultur og aggregatmetode: (i) Neuronal aggregering utføres i pyramidale mikrobrønn-arrays, støpt av 3D-trykte støper, som passer inn i brønnene på en 12-brønn kulturplate. (Ii) Mikro-TENNer inkluderer primære rotte-neuroner dissosiert fra føtalhjerner av embryonaldag-18-rotter. Etter vevsdissociasjon med trypsin-EDTA og DNase I fremstilles en celleoppløsning med en densitet på 1,0-2,0 x 106 celler / ml. (Iii) 12 μl av denne løsningen overføres til hver brønn i den pyramide mikrobrønn array. Platen som inneholder disse mikrobrønnene, blir sentrifugert for å produsere celleaggregater. (Iv) Disse inkuberes deretter over natten før plating i mikrokolonnene. ( C ) ECM-kjernefabrikasjon og cellesøing: (i) Før cellesøing ble en ECM-løsning inneholdende 1 mg / ml type I kollagen og 1 mg / mlLaminin overføres til det indre av mikro-tennene og får lov til å polymerisere. (Ii) Avhengig av om enveis eller toveis mikro-TENN er tilveiebragt, blir et aggregat plassert ved henholdsvis en eller begge ytterpunkter av mikrokolonnen. (Iii) Etter en inkuberingsperiode for å fremme vedheft blir mikro-TENN dyrket i petriskåler oversvømt med supplert embryonalt nevronbasalt medium. Iv) Etter 3-5 dager i kultur bør den endelige mikro-TENN-strukturen demonstrere celleaggregater ved ekstremer av mikrokolonnen, med aksonale kanaler som strekker seg over lengden. Vennligst klikk her for å se en større versjon av denne figuren.
