Two-step lithography: Figure 5 shows the result of a two-step lithographic process on a glass slide with overlapping patterns of fluorescently labeled DIS strands.
Expression of a fluorescent protein from a gene brush: Figure 6 demonstrates the expression of the fluorescent protein YPet from immobilized DNA. At several points in time we assessed the gene expression rate by partly bleaching the fluorescent protein and observing the recovery of the fluorescence signal, disregarding the immediate recovery, which does not result from protein expression. After the first bleaching at two hours of expression, the fluorescence intensity recovered quickly and rose above its value before the bleaching. After four and six hours, the fluorescence did not recover to its previous intensity, indicating that without the supply of fresh expression mix, the reaction terminated after approximately 3-4 h.
Coupling to microfluidics: Gene expression can be sustained over longer periods of time by supplying the expression compartments with additional precursor molecules via microfluidics. Figure 7 shows such a system, enabling the expression of YPet over 10 h.
TIRF observation: Bephore can also be applied to study the interaction of fluorescently labeled proteins with a DNA brush at the single molecule level. Especially in a noisy environment, lithography helps to distinguish between specific and unspecific interaction with the brush or the surface, respectively. Figure 8 gives such an example, with fluorescently labeled T7 RNA polymerase binding or adhering preferentially to the DNA brush compared to the surrounding surface.
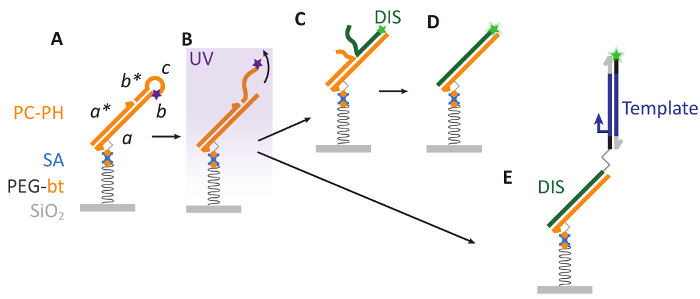
Figure 1: Bephore photolithography. A. A substrate with a surface of silicon dioxide (SiO2) is covered with a layer of biotinylated polyethylene glycol (PEG-bt), which is biocompatible and allows for the attachment of a photocleavable DNA hairpin via biotin-streptavidin interactions. Here, PC contains sequence domains abcb* and a photocleavable modification (purple star) and is hybridized to the strand PH with domain a*. B. Ultraviolet (UV) illumination cleaves PC, releasing a DNA fragment (cb*) into solution. C-D. The resulting single-stranded region on PC (b) aids as a toehold for the displacement of PH by a fluorescently labeled (green star) DIS strand. E. Also longer, double-stranded DNA ("Template") can be attached to the patterned surface. Such DNA is prepared by PCR with a primer carrying a DIS sequence at its 5' end, where primer and DIS are separated by a triethylene glycol spacer to keep DIS single-stranded during PCR (see also supplementary information, sections 2-4). Please click here to view a larger version of this figure.

Figure 2: Sample preparation for photolithography. A. Place a Bephore chip with an alignment mark on a microscopy slide or another chip holder (e.g. the holder in Figure 3B). Apply two-component silicone glue as a hydrophobic barrier along the edges of the chip (steps 3.1.2-3). B. Place the mask onto a mask holder. Here, we removed the iris of the field stop and modified its holder so the mask can be held by small magnets. For precise (angular) alignment in multi-step lithography, ensure that alignment marks on the holder and the mask match (step 3.2.1). C. To navigate on the chip and to align the mask with the alignment marks on the chip, slide in the red filter. For the UV exposure, insert the UV filter (steps 3.2.2-5). Figure reproduced from the Supporting Information of previous work27. Please click here to view a larger version of this figure.

Figure 3: Assembly of a holder for the observation of compartmentalized gene expression. A. Parts of the holder (ruler unit: cm). B-C. Top and bottom part of the holder are assembled separately, with the bottom holder carrying a patterned Bephore chip (section 5.1) and the top holder carrying a PDMS chip with compartments (section 5.2). D-F. Chip and PDMS are carefully brought into tight contact, while simultaneously observing and aligning compartments and DNA brushes in a stereoscopic microscope (steps 5.3.1-6). G. Before transferring the holder to an inverted, temperature-controlled microscope, the whole system is encapsulated in an anti-evaporation box (steps 5.3.7-8). Figure reproduced from the Supporting Information of previous work27. Please click here to view a larger version of this figure.

Figure 4: Microfluidic setup and sample holder for the compartmentalized gene expression. A. Parts of the sample holder, the PDMS device, the temperature-controlled microscope stage and the Bephore glass slide (ruler unit: cm). B. A microscopy slide carrying the PDMS with compartments is glued to the PDMS holder and exposed to oxygen plasma together with the PDMS in a plasma cleaner. The PDMS is then inserted into the top holder (steps 6.2.2-5). C. The cell-free expression system tube is connected to a pressure controller and placed on ice. The tubing (red dashed line in the inset) for the cell-free expression system is cooled by rubber tubing (blue dashed line) through which ice water is pumped by a peristaltic pump (section 6.1). D. The tubing is connected to the inlet position on the PDMS device. Another piece of tubing is connected to the outlet (steps 6.2.6-7). E. The top holder is placed on the microscope stage and carefully lowered towards the Bephore slide. The PDMS holder can still be moved in the x-y-plane to align the compartments in the PDMS with the gene brush on the Bephore chip. The wingnuts are used to press the PDMS onto the Bephore chip and fix the top holder to the microscope stage (steps 6.4.1-3). F. Cell-free expression system is pumped through the micro-channels in the PDMS and gene expression from DNA brushes can be monitored in epifluorescence microscopy (step 6.4.4). Please click here to view a larger version of this figure.
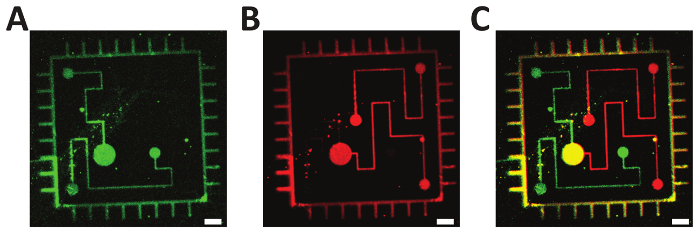
Figure 5: Two-step photolithography. A-B. Fluorescently labeled oligonucleotides (green: ATTO 532; red: Alexa Fluor 647) were consecutively immobilized on a Bephore glass slide via mask projection lithography with exposure times of 45 s. C. Overlay of subfigures A and B, demonstrating the precise alignment of the single exposures. Scale bars: 10 µm. Please click here to view a larger version of this figure.
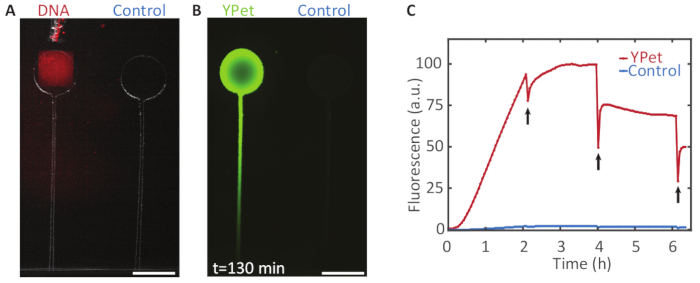
Figure 6: Compartmentalized gene expression. A. A DNA brush on a Bephore glass slide (UV exposure time: 2 min), coding for the fluorescent protein YPet was aligned with a compartment on a PDMS chip (see section 5 and Figure 3). B. Gene expression in the chamber with DNA yielded a strong fluorescence signal with a protein concentration gradient forming along a channel, which connected the chamber to the expression mix outside of the PDMS device. The control chamber without immobilized genes remained relatively dark. C. Fluorescence intensity profile over time for both chambers. Every two hours the fluorescent protein was partly bleached (black arrows) to check whether the expression was still active. After 4 h, the fluorescence did not recover to its previous intensity, indicating that expression had terminated. Scale bars: 300 µm. Please click here to view a larger version of this figure.
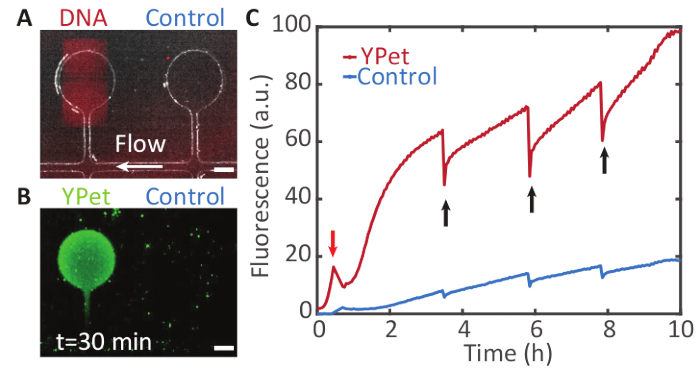
Figure 7: Sustained compartmentalized gene expression. A. A DNA brush on a Bephore glass slide (UV exposure time: 2 min), coding for YPet was aligned with a compartment on a PDMS chip. The compartments are connected to a supply channel (white arrow) through which cell-free expression system is pumped (see section 6 and Figure 4). B. The compartment containing the gene brush shows a fluorescence signal from YPet expression (in green) by the cell-free gene expression system. The neighboring compartment without a gene brush remains relatively dark. Fresh components of the cell-free expression system flow through the supply channel and diffuse into the compartments, while waste products are transported away. C. Fluorescence intensity profile over time for both chambers. The fluorescent proteins were partly bleached at different points in time (black arrows) to check whether expression was still active. Due to flow in the supply channel, gene expression was maintained for at least 10 h. (The peak in the red trace marked by the red arrow was caused by an air bubble that temporarily drained the solution from the compartments). Scale bars: 50 µm. Please click here to view a larger version of this figure.
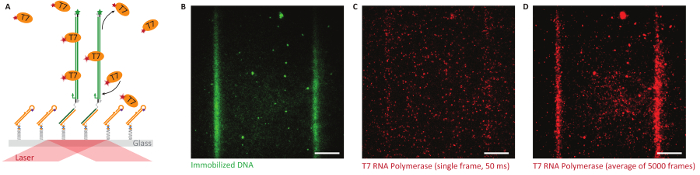
Figure 8: Single-molecule studies on Bephore glass slides in total internal reflection fluorescence microscopy (TIRFM). A. Objective-type TIRFM enables single-molecule imaging close to the glass-water interface. We immobilized fluorescently labeled genes (green, ATTO 532, UV exposure time: 2 min) with a T7 promoter along lithographically defined stripes and observed the behavior of T7 RNA polymerase (orange, labeled with Alexa Fluor 647) interacting with the surface. B. Fluorescence image showing two stripes of immobilized genes. C. T7 RNA polymerases attach specifically and non-specifically to the surface (single image, 50 ms exposure time). D. An average image obtained from all frames of a fluorescence video (5,000 frames like in subfigure C) exposes the specific interaction of the RNA polymerase with the DNA brush. Scale bars: 10 µm. Please click here to view a larger version of this figure.
Supplemental File 1. Please click here to download this file.
Supplemental File 2. Please click here to download this file.
