The representative results in Figure 1 and Figure 5 show data from two enzymes, lactase and invertase. Lactase and invertase catalyze the hydrolysis of a disaccharide into two monosaccharides, endothermically and exothermically, respectively. Both enzymatic reactions were run at concentrations that precluded saturation of the enzyme.
The lactase data demonstrate how ITC data can be used to estimate enzyme stability. Four sequential 4 µL injections of 600 mM lactose (Figure 1) were titrated into 20 mg/mL lactase. An interval of 5,400 s between each injection was applied, and this allows enough time to return to the initial baseline before each injection. The protocol described above was performed at 25 °C, 35 °C, 45 °C, and 55 °C. Additionally, 600 mM lactose was titrated into 100 mM sodium acetate buffer only at 55 °C as a control for the heat of mixing. For each injection of enzyme into substrate, there is an initial exothermic heat of mixing, and subsequently the lactase catalyzed endothermic hydrolysis of lactose occurs until the reaction is completed and the heat rate returns to the baseline. The injection of lactose into the lactase solution is then repeated three more times with the peak height of the endothermic reaction from each subsequent injection a little less than the preceding injection because of the decrease of enzymatic activity. The raw data can then be converted to show the peak heat relative to time (Figure 2A-D). The peak height for each injection can then be fit to a linear regression and the slope indicates enzyme stability at the chosen temperature. The more negative the slope, the less stable the enzyme. As expected, enzyme stability decreases with increasing temperature (Figure 2E).
Each injection of lactose described in Figure 1 results in dilution of lactase. To demonstrate that this dilution is not the cause of the decrease in enzymatic activity, the ITC lactase activity assay was also done at 25 °C, the lactase concentration at the first injection and at the last injection (Figure 3). This dilution results in an 8% decrease in enzyme activity, whereas the fourth injection in the assay shows a 73% decrease in activity. The dilution of the lactase during the four-injection experiment thus had a relatively small effect (i.e., 11%) on enzyme activity. The actual loss of activity was therefore 73-8 = 65%.
As mentioned in the introduction one of the advantages of using ITC is that these reactions can be done in opaque media, such as milk. To demonstrate this capability of ITC, milk was injected directly into lactase (Figure 4). Because the pH of the milk is not matched to the sodium acetate buffer, there is a large exothermic heat of mixing immediately following the injection. The exothermic heat of mixing peak is seen in the control that lacks lactase (Figure 4, gray line) and in the two injections with milk (Figure 4, black and dotted lines) subsequent to the heat of mixing peak. The endothermic reaction indicating lactase activity occurs. Milk contains a complex mixture of proteins, vitamins, minerals, and lactose. Because of the complexity of milk, it is probable that other reactions occur during the course of our reaction. To demonstrate that the endothermic peak is due to lactase activity, the milk was spiked with 146 mM lactose. In the reaction with the lactose spiked milk (Figure 4, dotted line), the endothermic peak and area under the curve are larger than in the milk alone (Figure 4, black line), indicating that the endothermic peak is indeed due to the lactase activity. The baseline offset indicates that a slow reaction continues after the lactose reaction is finished.
To demonstrate how this assay can be used to compare the stability of enzymatic activity of two different enzyme preparations, the stability of free invertase and invertase immobilized on a nylon-6 nanofiber membrane are compared in Figure 5 at 35 °C11. As in the lactase assay, four 4 µL injections of sucrose were made sequentially and the maximum heat rate determined from the peak height for each injection. As shown in Figure 6, the peak heights decrease linearly with time indicating decreasing enzyme activity.
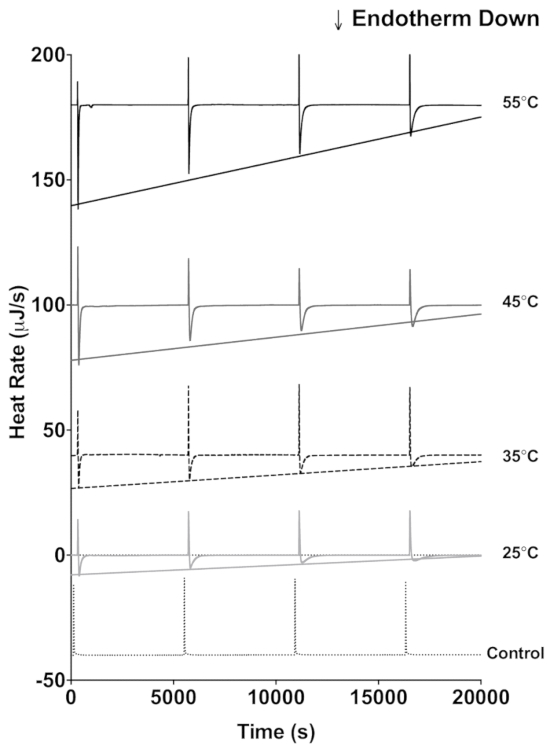
Figure 1: ITC data traces of lactase activity. Each trace shows four sequential 4 µL injections of 600 mM lactose into a lactase solution in pH 4.6 buffer. The traces were done at 55 °C (black), 45 °C (dark gray), 35 °C (dashed), 25 °C (light gray), and a no enzyme control (dotted) at 55 °C. The straight lines represent the fit to the endothermic peak minimum following each injection. Please click here to view a larger version of this figure.
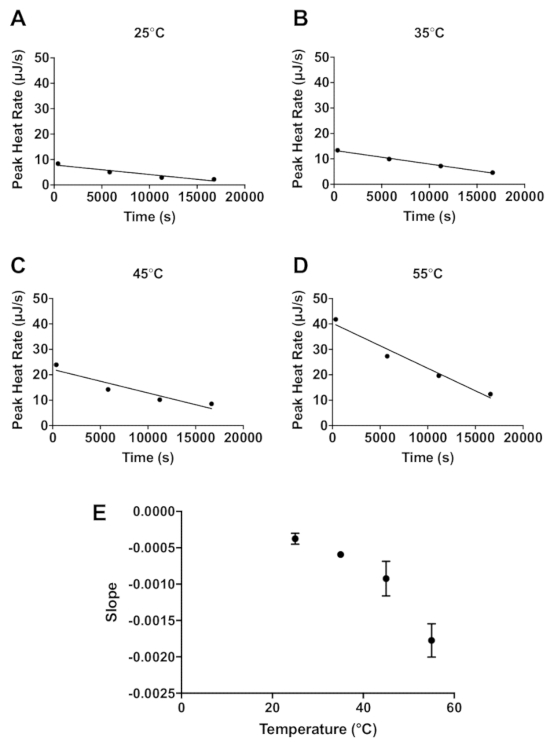
Figure 2: Stability of enzyme activity based on the rate of change in peak activity. The peak enzymatic activity for each of four injections relative to time (s) at 25 °C (A), 35 °C (B), 45 °C (C), and 55 °C (D). The linear fit of the data is represented by the solid line. The slopes of the fitted lines in parts A-D are plotted against temperature in E. Please click here to view a larger version of this figure.
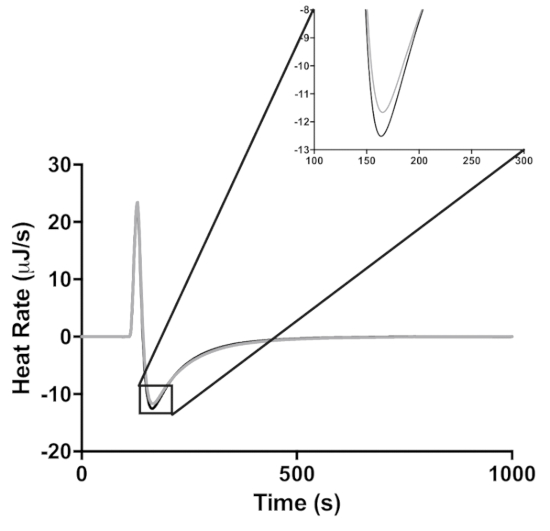
Figure 3: The effect of enzyme dilution on peak height at 25 °C. Lactase activity is measured at the enzyme concentration at the first and fourth injections of 20 mg/mL (black) and 18.62 mg/mL (gray), respectively. The inset is a zoomed in view of the peak enzyme activity. Please click here to view a larger version of this figure.
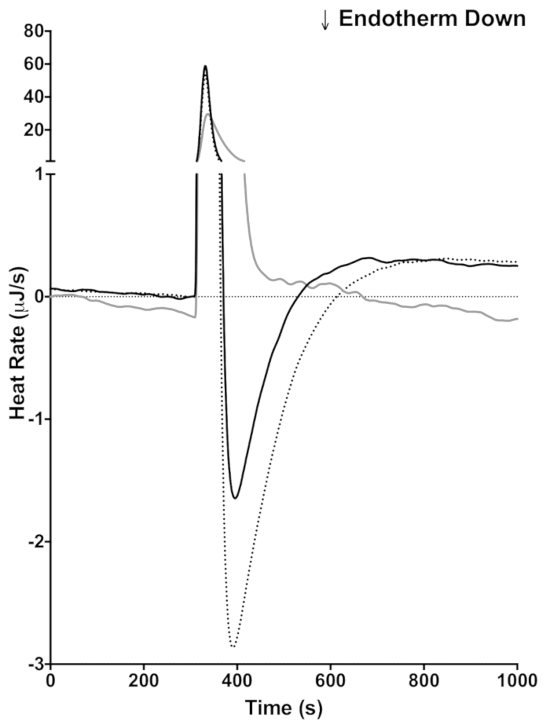
Figure 4: Measuring lactase activity in milk. ITC trace of an injection of BYU creamery fat free milk into buffer (gray), milk into 20 mg/mL lactase (black), and milk spiked with 5% additional lactose into 20 mg/mL (dotted). The y axis is discontinuous to show the endothermic enzyme reaction and the exothermic heat of mixing. Please click here to view a larger version of this figure.
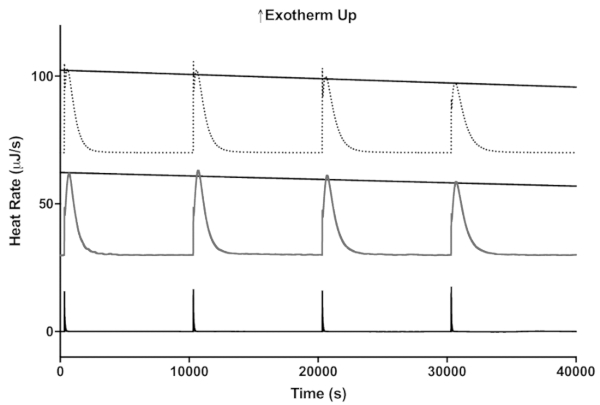
Figure 5: A representative example of how the ITC enzyme stability assay at 35 °C could be used to compare two different preparations of invertase. The activities of invertase immobilized on a nanofiber membrane and free enzyme are shown by the dotted and dark gray lines, respectively. The black line is the control with no invertase showing just the heat of mixing on appropriate scales. Please click here to view a larger version of this figure.
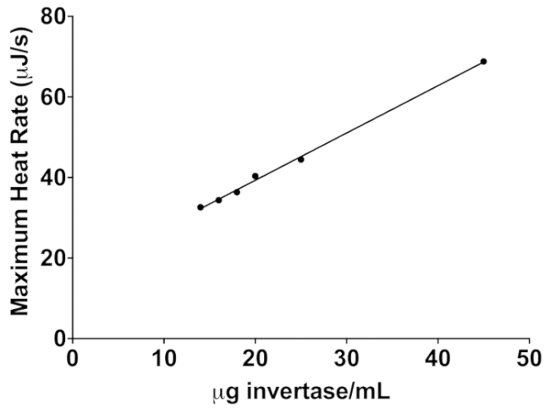
Figure 6: The linear relationship between peak exothermic heat rate and invertase concentration following a 4 µL injection of 600 mM sucrose. This standard curve can be used to determine the concentration of invertase in an unknown sample. Please click here to view a larger version of this figure.
