Reverse transkriptase Polymerasekædereaktionen (PCR) har været guldstandarden til mRNA-kvantificering. To assays, digital PCR (dPCR)1 og kvantitative, real time PCR (qPCR)2 bruges aktuelt. Af de to PCR teknikker har dPCR større følsomhed end qPCR tyder på, at det kunne bruges til at måle mRNA overflod i enkelte celler. Men i vores hænder, dPCR analyse af lav overflod mRNAs i puljer af 5 til 10 oocyter pr. hver eksperimentel prøve har produceret data med lav reproducerbarhed og høj variation3. Dette er sandsynligvis på grund af eksperimentelle fejl forbundet med RNA udvinding og reverse transkription effektivitet. RNA sekventering har også udført ved hjælp af en enkelt mus og menneskelige oocyter4,5. Denne teknik kræver cDNA forstærkning trin, der kræves for den bibliotek generation, hvilket sandsynligvis øger variation inden for en eksperimentel musikgruppe. Endvidere må lav overflod udskrifter ikke kunne påvises. Selvom sekventering priser er gået ned de sidste par år, kan det stadig være omkostningseffektivt uoverkommelige på grund af de høje udgifter til Bioinformatik analyser. Endelig, mRNA lokalisering er en dynamisk proces med rumlige ændringer bidrager til protein funktion6. Derfor, vi satte sig for at vedtage en teknik, der vil producere nøjagtige og reproducerbare kvantitative foranstaltninger og lokalisering af individuelle mRNAs i enkelt oocyter.
Forgrenede DNA koblet til fluorescens i situ hybridisering forstærker fluorescens signal snarere end forstærkende RNA/cDNA muliggør påvisning af enkelt mRNAs i enkelte celler 7,8,9. Analysen udføres gennem en række hybridisering, forstærkning (ved hjælp af forgrenede DNA) og fluorescens mærkning skridt for at forstærke fluorescens signal7. Teknikken, der begynder med bindingen af 18 – 25 base oligonukleotid sonde-par, der supplerer en specifik mRNA3,8,10. Femten til tyve sonde par er designet til hver afskrift sikring specificitet for target udskrift. MRNA-specifikke hybridisering er efterfulgt af pre-forstærker og forstærker sonder, der danner et forgrenet konfiguration. Ca, 400 etiket fluorophores binder til hver forstærker, hvilket resulterer i en 8000-fold stigning i fluorescens giver mulighed for påvisning af individuelle mRNAs (figur 1)11.
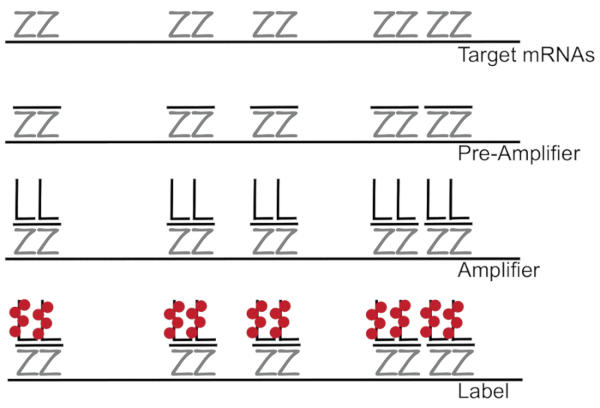
Figur 1: skematisk af SM-fisk protokol. Sekventiel hybridisering af udskrift specifikke sonde, forgrenet DNA forstærker og fluorophore til et mål mRNA er vist. Venligst klik her for at se en større version af dette tal.
Tidligere undersøgelser ved hjælp af enkelt molekyle fluorescens i situ hybridisering (SM-fisk) lokaliseret β-actin mRNAs i individuelle neuroner12 og humant papillomvirus DNA i livmoderhalskræft celle linjer7. Computersoftware stedet at finde og programmet til sporing af identificerer enkelte punktformet fluorescerende signal og held har været anvendt til at opgøre antallet af mRNAs i hver celle3,13.
Baseret på resultaterne af mRNA påvisning i neuroner12, hypotese vi at SM-fisk ville vise sig at være et nyttigt redskab til at kvantificere udskrift niveauer i murine oocytter og embryoner herunder lav overflod mRNAs. Men teknikken er optimeret til brug med vedhængende fast celler og formaldehyd fast paraffin indlejret (FFPE) væv sektioner. Oocyter kan ikke overholde et dias, selv når de er belagt med Poly-L-lysin. Derudover er de mere skrøbelige end somatiske celler og væv sektioner resulterer i celle lysis når de udsættes for nogle af leverandørejet bufferne i kommercielt tilgængelige kits3. For at overvinde disse udfordringer, oocyter fast og manuelt overført mellem dråber af buffere. Desuden blev permeabilization og vask buffere i kits udskiftet for at reducere celle lysering. Foruddesignede sonder er købt sammen med fisk kit eller specifikke udskrifter kan bestilles. Hver proprietære sonde sæt er tilgængelige i en af tre fluorescens kanaler (C1, C2 og C3) at tillade multiplexing. I den aktuelle eksperiment var murine oocyter dual-farvede og kvantificeret ved hjælp af en C2 Nanog sonde og en C3 Pou5f1 sonde. Disse prober blev udvalgt på baggrund af de rapporterede udtryk for Nanog og Pou5f1 i oocytter og embryoner. Ved afslutningen af hybridisering trin blev oocyter placeret i dråber af anti-fade montering medier for ansøgning til histologisk dias. Konfokal billeder blev brugt til at opgøre antallet af punktformet fluorescerende signaler, som repræsenterer individuelle mRNAs. Ud over at kvantificere mRNAs, imaging viste også den geografiske fordeling af den specifikke mRNA i cellen, er som andre RNA kvantificering metoder ude af stand til at opnå. Denne teknik vist sig for at have lav variation inden for en eksperimentel musikgruppe der tillader anvendelse af mindre antal oocyter i hver eksperimentelle gruppe for at identificere væsentlige forskelle mellem eksperimentelle grupper3.
