Reverse-transcriptase polymerase-kettingreactie (PCR) is de gouden standaard voor mRNA kwantificatie. Twee tests, digitale PCR (dPCR)1 en kwantitatieve, real time PCR (qPCR)2 worden momenteel gebruikt. Van de twee PCR technieken heeft dPCR grotere gevoeligheid dan qPCR suggereren dat het kan worden gebruikt voor het meten van de mRNA overvloed in afzonderlijke cellen. Echter, in onze handen, dPCR analyse van lage overvloed mRNAs in poules van 5 tot en met 10 eicellen per elk experimentele monster heeft gegevens met lage reproduceerbaarheid en hoge variatie3. Dit is waarschijnlijk te wijten aan de experimentele fout RNA extractie en omgekeerde transcriptie efficiëntie is gekoppeld. RNA sequencing is ook uitgevoerd met behulp van een enkele muis en menselijke eicellen4,5. Deze techniek vereist cDNA amplificatie stappen die nodig zijn voor de bibliotheek-generatie waardoor waarschijnlijk variabiliteit binnen een experimentele groep. Bovendien, lage overvloed afschriften mogelijk niet aantoonbaar. Hoewel sequencing prijzen zijn gedaald in de afgelopen jaren, kan het nog steeds kosten onbetaalbaar vanwege de hoge kosten van bioinformatica analyses. Ten slotte, mRNA lokalisatie is een dynamisch proces met ruimtelijke veranderingen bij te dragen aan eiwit functie6. Daarom zetten we uit te nemen van een techniek die nauwkeurige en reproduceerbare kwantitatieve maatregelen en lokalisatie van individuele mRNAs in enkele eicellen produceren zou.
Vertakte DNA gekoppeld aan fluorescentie in situ hybridisatie versterkt fluorescentie signaal in plaats van uitdeinende RNA/cDNA inschakelen detectie van één mRNAs in afzonderlijke cellen 7,8,9. De test wordt uitgevoerd door middel van een reeks van hybridisatie, versterking (met behulp van vertakte DNA) en fluorescentie labeling stappen om het versterken van de fluorescentie signaal7. De techniek begint met bindende 18 – tot 25-base oligonucleotide sonde-paren die complementair aan een specifieke mRNA3,8,10 zijn. Vijftien tot twintig sonde paren zijn ontworpen voor elke specificiteit transcript te zorgen voor het transcript van de doelgroep. De mRNA specifieke hybridisatie wordt gevolgd door voorversterker en versterker sondes die een vertakte configuratie vormen. Ongeveer binden 400 label fluorophores aan elke versterker, resulterend in een verhoging van de 8000-fold in fluorescentie waardoor detectie van individuele mRNAs (Figuur 1)11.
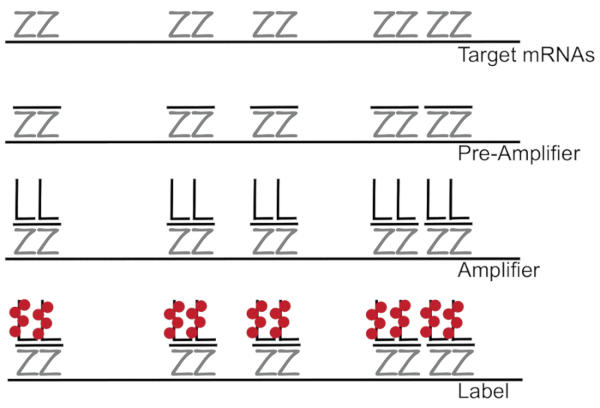
Figuur 1: schematische van de SM-vis protocol. Sequentiële kruising van transcript specifieke sonde, vertakt DNA versterker en fluorophore naar een doel dat mRNA wordt weergegeven. Klik hier voor een grotere versie van dit cijfer.
Eerdere studies met behulp van één molecuul fluorescentie in situ hybridisatie (SM-vis) gelokaliseerd β-actine mRNAs in individuele neuronen12 en humaan papillomavirus DNA in cervicale kanker cel lijnen7. De computersoftware plek vinden en Tracking programma identificeert individuele punctate fluorescent signaal en is met succes gebruikt om het aantal mRNAs in elke cel3,13te kwantificeren.
Op basis van de resultaten van mRNA detectie in neuronen12, veronderstelde wij dat SM-vis zou blijken een nuttig hulpmiddel om te kwantificeren transcript niveaus in lymfkliertest eicellen en embryo’s met inbegrip van lage overvloed mRNAs. Echter, de techniek is geoptimaliseerd voor het gebruik met aanhangend vaste cellen en formaldehyde vaste paraffine ingesloten weefselsecties (FFPE). Eicellen niet kunnen voldoen aan een dia, zelfs wanneer ze zijn bekleed met een Poly-L-lysine. Bovendien zijn ze kwetsbaarder dan somatische cellen en weefselsecties resulterend in lysis van de cel wanneer onderworpen aan sommige van de merkgebonden buffers in commercieel beschikbare kits3. Om deze uitdagingen te overwinnen, waren de eicellen vast en handmatig worden overgedragen tussen de buffers druppels. Bovendien, permeabilization en wastafel buffers in de kits werden vervangen ter vermindering van de lysis van de cel. Vooraf ontworpen sondes worden gekocht naast de vis kit of specifieke afschriften kunnen aangevraagd worden. Elke set merkgebonden sonde is beschikbaar in een van de drie kanalen van de fluorescentie (C1, C2 en C3) om multiplexing mogelijk te maken. In het huidige experiment waren lymfkliertest eicellen dual-gekleurd en gekwantificeerde met behulp van een sonde C2 Nanog en een C3 Pou5f1 -sonde. Deze sondes werden geselecteerd op basis van de gerapporteerde uitdrukking van Nanog en Pou5f1 in eicellen en embryo’s. Aan het einde van de kruising stappen, werden de eicellen in druppels anti-fade montage media voor toepassing op histologische dia’s geplaatst. Confocale afbeeldingen werden gebruikt om het kwantificeren van het aantal punctate fluorescerende signalen die individuele mRNAs vertegenwoordigen. Naast het kwantificeren van de mRNAs, imaging bleek ook de ruimtelijke spreiding van de specifieke mRNA in de cel, zijn welke andere RNA kwantificering methoden niet in staat om te bereiken. Deze techniek bleek te hebben lage variabiliteit binnen een experimentele groep toestaan van het gebruik van kleinere aantallen van eicellen in elke experimentele groep te identificeren van significante verschillen tussen experimentele groepen3.
