Обратной транскриптазы полимеразной цепной реакции (ПЦР) был золотым стандартом для мРНК quantitation. В настоящее время используются два анализов, цифровой ПЦР (dPCR)1 и количественных, реальное время ПЦР (ПЦР)2 . Из двух методов ПЦР dPCR имеет большую чувствительность, чем ПЦР предполагая, что он может использоваться для измерения mRNA изобилия в одиночных клетках. Однако в наших руках, dPCR анализ низкого залегания mRNAs в бассейнах 5 до 10 яйцеклеток за каждый экспериментальный образец выпустила данных с низкой воспроизводимостью и высокий вариант3. Это, вероятно, из-за экспериментальной ошибки, связанной с РНК добыча и эффективности обратной транскрипции. РНК последовательности также была выполнена с помощью мыши и человеческих яйцеклеток4,5. Этот метод требует cDNA амплификации шаги, необходимые для библиотеки поколения, которое вероятно увеличивает изменчивость в экспериментальной группе. Кроме того низкие изобилии стенограммы не может быть обнаружено. Хотя последовательности цены снизились за последние несколько лет, это может быть дорого из-за высокой стоимости биоинформатики анализов. Наконец мРНК Локализация — это динамичный процесс с пространственных изменений, способствующих белков функции6. Таким образом мы намереваемся принять метод, который будет производить точные и воспроизводимые количественные показатели и локализации индивидуальных mRNAs в одно ооцитов.
Разветвленный ДНК, в сочетании с флуоресценции в гибридизации situ усиливает сигнал флуоресценции, вместо того, чтобы усилительных благоприятных обнаружение РНК/кДНК одной мРНК в отдельных клетках 7,8,9. Генотипирования через серию гибридизации, амплификация (с помощью разветвленной ДНК) и флуоресценции маркировки шаги для того, чтобы усилить флуоресценции сигнала7. Этот метод начинается с привязки 18 – 25-base олигонуклеотида зонд пар, которые дополняют конкретные мРНК3,8,10. Пятнадцати до двадцати зонд пары предназначены для обеспечение специфику каждого Стенограмма для целевого транскрипт. МРНК конкретных гибридизации следуют предварительный усилитель и усилитель зондами, которые формируют разветвленную конфигурации. Приблизительно 400 лейбл флуорофоров привязку каждого усилителя, что привело к увеличению 8000-fold в флуоресцировании, позволяя обнаружения индивидуальных mRNAs (рис. 1)11.
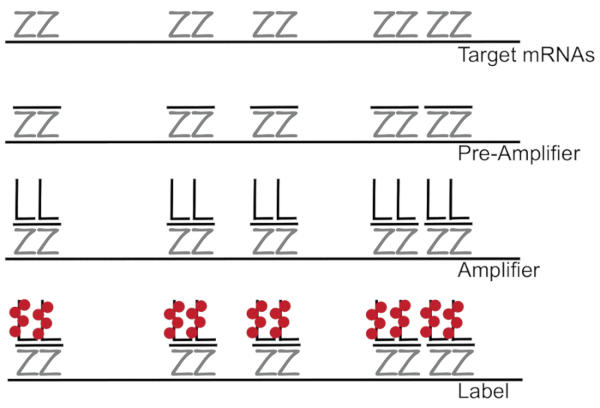
Рисунок 1: Схема протокола SM-рыба. Последовательные гибридизации Стенограмма конкретного ПЭП, разветвленной ДНК усилитель и Флюорофор к цели, которую показана mRNA. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
Предыдущие исследования с использованием одной молекулы флуоресценции в situ гибридизация (SM-рыбы) локализованные β-актина mRNAs в12 отдельных нейронов и ДНК вируса папилломы человека в рак шейки матки клеток линии7. Компьютерное программное обеспечение, месте нахождения и отслеживания Программа идентифицирует отдельных пунктата флуоресцентного сигнала и успешно используется для количественного определения количество мРНК в каждой ячейке3,13.
Основываясь на результаты обнаружения мРНК в нейроны12, мы предположили, что SM-рыб окажется полезным инструментом для quantitate Стенограмма уровней в мышиных яйцеклеток и эмбрионов, включая низкие изобилии мРНК. Однако техника оптимизирована для использования с фиксированной адэрентных клеток и формальдегида фиксированной парафин встроенных разделов ткани (FFPE). Ооциты не может присоединиться к слайду, даже когда они покрыты с поли-L-лизин. Кроме того они являются более хрупкие, чем соматических клетках и результате лизис клеток когда подвергаются некоторые несвободные буферов в коммерчески доступные комплекты3разделах ткани. Для преодоления этих проблем, ооциты фиксировали и вручную переведен между капли буферов. Кроме того permeabilization и мыть буферов в наборы были заменены уменьшить lysis клетки. Встроенные датчики приобретаются вместе с комплектом рыбу или конкретных протоколов могут запрашиваться. Каждого собственности зонда набор доступен в одном из трех каналов флуоресцирования (C1, C2 и C3) для мультиплексирования. В ходе текущего эксперимента мышиных ооциты были двойной окрашенных и количественных, используя зонд Nanog C2 и C3 Pou5f1 зонда. Эти зонды были отобраны на основе сообщенных выражения Nanog и Pou5f1 яйцеклеток и эмбрионов. В заключение гибридизации шаги ооциты были помещены в капли анти затухания монтажа СМИ для приложения гистологические слайды. Конфокальный изображения были использованы для количественного определения количество пунктата флуоресцентные сигналов, которые представляют индивидуальных mRNAs. Помимо количественного мРНК, изображений также показал пространственное распределение конкретные мРНК в ячейке, какие другие методы количественного определения РНК не в состоянии достичь. Этот метод оказался низким изменчивости в пределах экспериментальной группы, что позволяет использовать меньшее количество яйцеклеток в каждой экспериментальной группе выявить существенные различия между экспериментальной группы3.
