Transcriptase reversa cadeia da polimerase (PCR) tem sido o padrão-ouro para quantificação de mRNA. Dois ensaios, digital PCR (dPCR)1 e quantitativo, real time PCR (qPCR)2 são utilizados atualmente. Das duas técnicas de PCR, dPCR tem maior sensibilidade do que qPCR, sugerindo que poderia ser usado para medir a abundância de mRNA em células únicas. No entanto, em nossas mãos, dPCR análise de mRNAs baixa abundância nas piscinas de ovócitos de 5 a 10 por cada amostra experimental produziu dados com baixa reprodutibilidade e alta variação3. Isto é provavelmente devido ao erro experimental associado a extração do RNA e transcrição reversa eficiência. A sequenciação do ARN também foi executada usando um único rato e oócitos humanos4,5. Esta técnica requer etapas de amplificação do cDNA necessárias para a geração de biblioteca que provavelmente aumenta a variabilidade dentro de um grupo experimental. Além disso, transcrições de baixa abundância podem não ser detectáveis. Embora os preços de sequenciamento caíram nos últimos anos, ainda pode ser custo proibitivo devido ao alto custo das análises de Bioinformática. Finalmente, a localização de mRNA é um processo dinâmico, com alterações espaciais, contribuindo para a proteína função6. Portanto, decidimos para adotar uma técnica que produziria medidas quantitativas precisas e reprodutíveis e localização dos mRNAs individuais em oócitos único.
DNA em cadeia ramificada acoplado a fluorescência hibridação in situ amplifica o sinal de fluorescência em vez de amplificação do RNA/cDNA permita deteção dos mRNAs único em células individuais 7,8,9. O ensaio é realizado através de uma série de hibridização e amplificação (usando DNA em cadeia ramificada) fluorescência rotulagem passos a fim de amplificar o sinal de fluorescência7. A técnica começa com ligação de pares de sonda de 18 a 25-base do oligonucleotide que são complementares a uma específica do mRNA3,8,10. Quinze a vinte pares de sonda são projetados para cada especificidade de transcrição garantindo para a transcrição do alvo. A hibridação de mRNA específico é seguida por sondas pré-amplificador e amplificador que formam uma configuração ramificada. Aproximadamente, 400 rótulo fluorophores bind para cada amplificador, resultando em um 8000-fold aumento na fluorescência permitindo a deteção de mRNAs individuais (Figura 1)11.
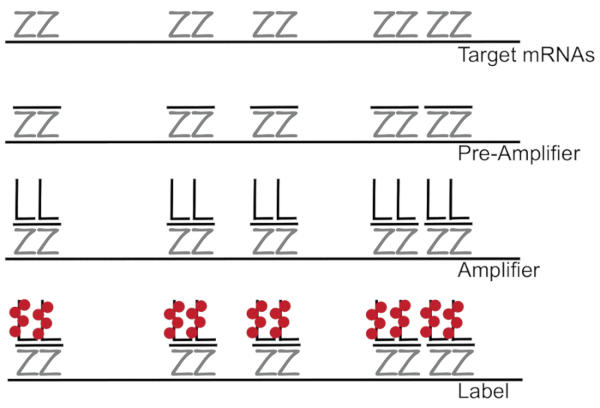
Figura 1: esquemático do protocolo SM-peixes. Sequencial hibridização da sonda específica de transcrição, ramificado DNA amplificador e fluoróforo para um destino de mRNA é mostrado. Clique aqui para ver uma versão maior desta figura.
Estudos anteriores usando fluorescência única molécula em situ da hibridação (SM-peixe) localizada β-actina mRNAs em neurônios individuais12 e DNA de papilomavírus humano em câncer de colo uterino de células linhas7. O software de computador, encontrar o local e o programa de rastreamento identifica o sinal fluorescente punctate individual e tem sido usada com sucesso para quantificar o número de mRNAs em cada célula3,13.
Baseado nos resultados da deteção do mRNA em neurônios12, formulamos a hipótese que SM-peixe provaria uma ferramenta útil para dosar os níveis de transcrição de murino oócitos e embriões, incluindo baixa abundância mRNAs. No entanto, a técnica é otimizada para uso com pilhas fixos aderentes e formaldeído fixada parafina incorporado cortes de tecido (FFPE). Oócitos não podem aderir a um slide, mesmo quando eles são revestidos com poli-L-lisina. Além disso, eles são mais frágeis do que as células somáticas e cortes de tecido, resultando em lise celular quando submetido a alguns dos buffers de proprietários em kits comercialmente disponíveis3. Para superar estes desafios, oócitos foram fixo e manualmente transferidos entre gotas dos buffers. Além disso, os buffers de permeabilização e lavagem nos kits foram substituídos para reduzir o lysis da pilha. Sondas preconcebidas são compradas juntamente com o kit de peixe ou transcrições específicas podem ser solicitadas. Cada conjunto de sonda proprietário está disponível em um dos três canais de fluorescência (C1, C2 e C3) para permitir a multiplexação. Na experiência atual, murino oócitos foram manchadas de dual e quantificados usando uma sonda de C2 Nanog e uma sonda de C3 Pou5f1 . Estas sondas foram selecionadas com base em expressão relatado de Nanog e Pou5f1 de oócitos e embriões. Na conclusão das etapas da hibridação, oócitos foram colocados em gotas de mídia anti-desvaneça-se montagem para aplicação de lâminas histológicas. Confocal imagens foram usadas para quantificar o número de sinais fluorescentes punctate que representam os mRNAs individuais. Além de quantificar os mRNAs, imagem também mostrou a distribuição espacial do mRNA específico na célula, quais outros métodos de quantificação de RNA são incapazes de alcançar. Esta técnica provou ter baixa variabilidade dentro de um grupo experimental, permitindo o uso de um menor número de oócitos em cada grupo experimental para identificar diferenças significativas entre os grupos experimentais3.
