Reazione a catena d’inversione-transcriptase della polimerasi (PCR) è stato il gold standard per la quantificazione di mRNA. Due saggi, digitale PCR (dPCR)1 e quantitativa in tempo reale PCR (qPCR)2 sono attualmente utilizzati. Le due tecniche di PCR, dPCR ha una maggiore sensibilità rispetto qPCR suggerendo che potrebbe essere utilizzato per misurare l’abbondanza del mRNA in singole cellule. Tuttavia, nelle nostre mani, dPCR analisi dei mRNAs abbondanza bassa nelle piscine di ovociti di 5 a 10 per ogni campione sperimentale ha prodotto dati con scarsa riproducibilità e variazione alta3. Ciò è probabilmente dovuto l’errore sperimentale associata con estrazione del RNA e l’efficienza di trascrizione inversa. Sequenziamento di RNA è stata eseguita anche utilizzando un unico mouse e ovociti umani4,5. Questa tecnica richiede passaggi di amplificazione del cDNA necessari per la generazione della libreria che probabilmente aumenta la variabilità all’interno di un gruppo sperimentale. Inoltre, trascrizioni di abbondanza bassa possono non essere rilevabile. Anche se i prezzi di sequenziamento sono scesi di ultimi anni, può ancora essere costi proibitivi a causa del costo elevato di analisi bioinformatica. Infine, la localizzazione di mRNA è un processo dinamico con cambiamenti spaziali contribuendo alla proteina funzione6. Pertanto, abbiamo deciso di adottare una tecnica che avrebbe prodotto accurate e riproducibile misure quantitative e localizzazione di singoli mRNA in singoli ovociti.
DNA ramificato accoppiato all’ibridazione fluorescente in situ amplifica il segnale di fluorescenza piuttosto che amplificazione RNA/cDNA abilitazione individuazione dei singoli mRNAs in singole celle 7,8,9. Il dosaggio è effettuato attraverso una serie di ibridazione, amplificazione (usando il DNA ramificato) e fluorescenza etichettatura passi al fine di amplificare il segnale di fluorescenza7. La tecnica comincia con l’associazione di coppie di sonda di 18 – 25-base del oligonucleotide che sono complementari a un specifico mRNA3,8,10. Quindici-venti coppie di sonda sono progettate per ogni specificità assicurando di trascrizione per la trascrizione di destinazione. L’ibridazione di specifici mRNA è seguita dalle sonde preamplificatore e amplificatore che formano una configurazione ramificata. Circa, 400 etichetta fluorofori associare a ciascun amplificatore, conseguente 8000-fold aumento in fluorescenza permettendo la rilevazione di singoli mRNA (Figura 1)11.
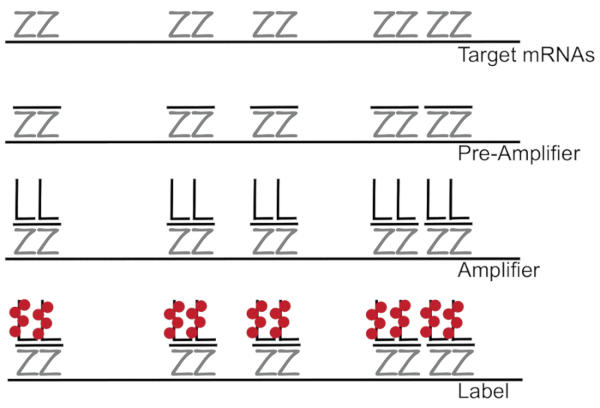
Figura 1: schema del protocollo SM-pesce. Sequenza ibridazione della sonda specifica trascrizione, ramificata amplificatore DNA e fluoroforo ad un target di mRNA viene mostrato. Clicca qui per visualizzare una versione più grande di questa figura.
Gli studi precedenti usando la singola molecola fluorescenza in situ di ibridazione (SM-pesce) localizzato β-actina mRNA in singoli neuroni12 e papillomavirus umano DNA nel cancro cervicale delle cellule linee7. Il software per computer trovare posto e programma di controllo identifica singolo segnale fluorescente punctato ed è stato usato con successo per quantificare il numero dei mRNAs in ogni cella3,13.
Sulla base dei risultati della rilevazione di mRNA in neuroni12, abbiamo supposto che SM-pesce si sarebbe rivelato un utile strumento per quantificare i livelli di trascrizione in murini ovociti ed embrioni tra cui abbondanza bassa mRNAs. Tuttavia, la tecnica è ottimizzata per l’utilizzo con celle fisse aderenti e formaldeide fissata paraffina embedded sezioni di tessuto (FFPE). Gli ovociti non possono aderire a una diapositiva, anche quando essi sono rivestiti con poli-L-lisina. Inoltre, essi sono più fragili di cellule somatiche e sezioni di tessuto con conseguente lisi cellulare quando sottoposto ad alcuni dei buffer proprietarie in kit commercialmente disponibili3. Per superare queste sfide, ovociti erano fissi e trasferiti manualmente tra gocce dei buffer. Inoltre, la permeabilizzazione e wash buffer nei kit sono stati sostituiti per ridurre la lisi delle cellule. Sonde predefiniti vengono acquistati a fianco il kit di pesce o possono essere richiesti specifici trascritti. Ogni set di sonde proprietarie è disponibile in uno dei tre canali di fluorescenza (C1, C2 e C3) per consentire per multiplexing. Nell’esperimento attuale, ovociti murini sono stati doppia colorazione e quantificati mediante una sonda di C2 Nanog e una sonda di C3 Pou5f1 . Queste sonde sono state selezionate basato sull’espressione di Nanog e Pou5f1 segnalato di ovociti ed embrioni. A conclusione delle operazioni di ibridazione, gli ovociti sono stati collocati in gocce di mezzi di montaggio anti-dissolvenza per vetrini istologici dell’applicazione. Immagini confocal sono stati utilizzati per quantificare il numero di segnali fluorescenti punctati che rappresentano singoli mRNA. Oltre a quantificare i mRNAs, formazione immagine ha mostrato anche la distribuzione spaziale di specifici mRNA nella cellula, quali altri metodi di quantificazione di RNA sono in grado di raggiungere. Questa tecnica ha dimostrato di avere bassa variabilità all’interno di un gruppo sperimentale che consente di utilizzare numeri più piccoli degli ovociti in ogni gruppo sperimentale per identificare differenze significative tra i gruppi sperimentali3.
