Reacción en cadena reversa-transcriptase de polimerasa (PCR) ha sido el estándar de oro para la cuantificación del mRNA. Actualmente se utilizan dos ensayos PCR (dPCR) digital1 y cuantitativa, real time PCR (qPCR)2 . De las dos técnicas PCR, dPCR tiene mayor sensibilidad que la qPCR sugiriendo que podría ser utilizado para medir la abundancia de ARNm en células individuales. Sin embargo, en nuestras manos, análisis de dPCR de mRNAs de baja abundancia en grupos de 5 a 10 ovocitos por cada muestra experimental ha producido datos con baja reproducibilidad y alta variación3. Esto es probablemente debido al error experimental asociado a la extracción de RNA y transcripción reversa eficiencia. La secuencia de RNA también se ha realizado utilizando un único ratón y ovocitos humanos4,5. Esta técnica requiere pasos de amplificación de cDNA para la generación de biblioteca que probablemente aumenta la variabilidad dentro de un grupo experimental. Además, las transcripciones de baja abundancia no pueden ser detectables. Aunque los precios de la secuencia han bajado en los últimos años, todavía puede ser prohibitivo debido al costo alto de Bioinformática análisis de costo. Por último, la localización de mRNA es un proceso dinámico con cambios espaciales que contribuyen a la función de la proteína6. Por lo tanto, nos propusimos adoptar una técnica que produciría medidas cuantitativas precisas y reproducibles y localización de los mRNAs individuales en ovocitos solo.
DNA ramificado juntada a hibridación fluorescente in situ amplifica la señal de fluorescencia en lugar de amplificar RNA/cDNA que permite detección de mRNAs individuales en células individuales 7,8,9. El ensayo se realiza a través de una serie de hibridación, amplificación (usando ADN ramificado) y fluorescencia de etiquetado pasos para amplificar la señal de la fluorescencia del7. La técnica comienza con el atascamiento de pares de sonda de 18 – 25 base de oligonucleótidos que son complementarias a un ARNm específico3,8,10. Quince a veinte pares de sonda están diseñados para cada especificidad asegurando de transcripción para la transcripción de destino. La hibridación específica del mRNA es seguida por sondas de preamplificador y amplificador que forman una configuración ramificada. Aproximadamente, 400 etiqueta fluoróforos se unen a cada amplificador, resultando en un 8000-fold incremento de fluorescencia que permite la detección de mRNAs individuales (figura 1)11.
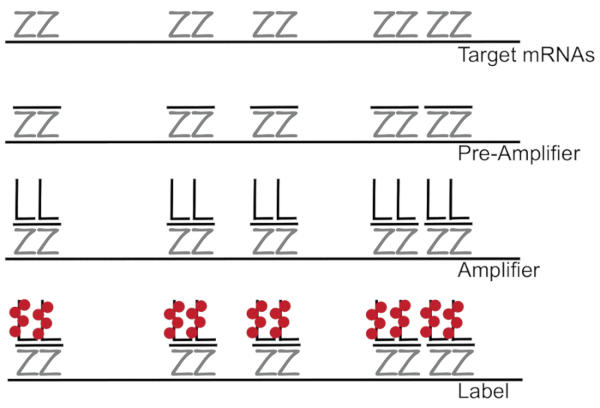
Figura 1: esquema del protocolo SM-pescado. Hibridación secuencial de sonda específica de la transcripción, ramificado ADN amplificador y fluoróforo a un mRNA se muestra de destino. Haga clic aquí para ver una versión más grande de esta figura.
Estudios previos usando in situ hibridación (SM-pescado) localizado β-actina mRNAs en neuronas individuales12 y papillomavirus humano DNA en cáncer de cuello uterino fluorescencia sola molécula célula líneas7. El software de computadora encontrar lugar y programa de seguimiento identifica la señal fluorescente punteada individual y se ha utilizado con éxito para cuantificar el número de mRNAs en cada célula3,13.
Basado en los resultados de la detección de mRNA en las neuronas12, presumimos que SM-pez sería una herramienta útil para cuantificar los niveles de transcripción en murinos ovocitos y embriones incluyendo mRNAs de baja abundancia. Sin embargo, la técnica está optimizada para el uso con las células fijas adherentes y formaldehído fijada parafina incorporado secciones de tejido (FFPE). Ovocitos no se adhieren a un portaobjetos, incluso cuando están recubiertos con poli-l-lisina. Además, son más frágiles que las células somáticas y las secciones de tejido que resulta en lisis celular cuando se someten a algunos de los buffers propietarios en kits disponibles en el mercado3. Para superar estos desafíos, ovocitos fueron fijos y transferidos manualmente entre las gotas de los amortiguadores. Además, buffers de permeabilización y lavado en los kits fueron substituidos para reducir la lisis celular. Sondas prediseñadas se compran junto con el kit de peces o pueden solicitar transcripciones específicas. Cada conjunto de sonda patentada está disponible en uno de los tres canales de fluorescencia (C1, C2 y C3) para permitir la multiplexación. En el experimento actual, oocitos murinos fueron cuantificados utilizando una sonda C2 Nanog y una sonda de C3 Pou5f1 y doble tinción. Estas sondas se seleccionaron con base en la expresión divulgada de Nanog y Pou5f1 en ovocitos y embriones. En la conclusión de los pasos de hibridación, ovocitos fueron colocados en gotas anti-fade del medio de montaje para aplicación a las diapositivas histológicas. Imágenes confocales fueron utilizados para cuantificar el número de señales fluorescentes punteadas que representan los mRNAs. Además de cuantificar los mRNAs, proyección de imagen demostró también la distribución espacial de los mRNA específicas en la célula, que otros métodos de cuantificación de RNA son incapaces de lograr. Esta técnica demostró para tener poca variabilidad dentro de un grupo experimental que permite el uso de un número menor de ovocitos en cada grupo experimental para identificar diferencias significativas entre grupos experimentales3.
