Revers transkriptase polymerasekjedereaksjons (PCR) har vært gullstandarden for mRNA kvantifisering. To analyser, digital PCR (dPCR)1 og kvantitativ, virkelig tid PCR (qPCR)2 brukes for øyeblikket. Av de to PCR teknikkene har dPCR større følsomhet enn qPCR antyder at det kan brukes til å måle mRNA overflod i enkeltceller. Men i våre hender, har dPCR analyse av lav overflod mRNAs i av 5 til 10 oocytes per hvert eksperimentelle utvalg produsert data med lav reproduserbarhet og høy variant3. Dette er sannsynligvis på grunn av eksperimentelle feil forbundet med RNA utvinning og omvendt transkripsjon effektivitet. RNA sekvensering er også utført med et enkelt museklikk og menneskelig oocytes4,5. Denne teknikken krever cDNA forsterkning fremgangsmåtene for biblioteket generasjon som sikkert øker variasjonen i en forsøksgruppen. Videre kan lav overflod utskrifter ikke være synlig. Selv om sekvensering prisene har gått ned de siste årene, kan det likevel være kostnadseffektivt uoverkommelige på grunn av de høye kostnadene av bioinformatikk analyser. Endelig er mRNA lokalisering en dynamisk prosess med romlig endringer bidra til protein funksjon6. Derfor setter vi opp for vedta en teknikk som vil produsere nøyaktig og reproduserbar kvantitative tiltak og lokalisering av personlige mRNAs i én oocytes.
Forgrenede DNA koblet til fluorescens i situ hybridisering forsterker fluorescens signal i stedet for å forsterke RNA/cDNA aktivere gjenkjenning av ett mRNAs i enkeltceller 7,8,9. Analysen utføres gjennom en rekke hybridisering, forsterkning (med forgrenet DNA) og fluorescens merking skritt for å forsterke fluorescens signal7. Teknikken begynner med binding av 18 – til 25-base oligonucleotide sonde par som er komplementære til en bestemt mRNA3,8,10. Femten til tjue sonde parene er designet for hver utskrift sikre spesifisitet for målet transkripsjon. MRNA-spesifikke hybridization etterfølges av pre forsterker og forsterker sonder som danner en forgrenet konfigurasjon. Ca, 400 etiketten fluorophores binder seg til hver forsterker, resulterer i en 8000-fold økning i fluorescens tillater påvisning av individuelle mRNAs (figur 1)11.
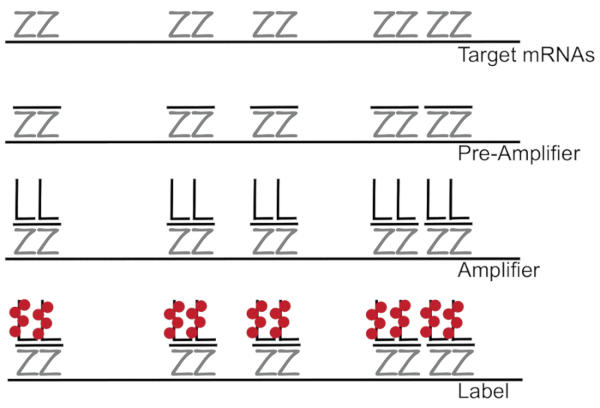
Figur 1: skjematisk av SM-fisk protokollen. Sekvensiell blanding av transkripsjon bestemt sonde, forgrenet DNA forsterker og fluorophore til et mål mRNA vises. Klikk her for å se en større versjon av dette tallet.
Tidligere studier med ett molekyl fluorescens i situ hybridisering (SM-fisk) lokalisert β-utgangen mRNAs i individuelle neurons12 og humant papillomavirus DNA i livmorhalskreft celle linjer7. Dataprogramvaren sted å finne og sporing programmet identifiserer individuelle vises punctate fluorescerende signal og har blitt brukt å tallfeste antallet mRNAs i hver celle3,13.
Basert på resultatene av mRNA gjenkjenning i neurons12, hypotese vi at SM-fisk ville bevise en nyttig verktøyet å quantitate transkripsjon nivåer i murine oocytes og embryo inkludert lav overflod mRNAs. Men teknikken er optimalisert for bruk med tilhenger fast celler og formaldehyd fast parafin innebygd (FFPE) vev deler. Oocytes kan ikke følge et lysbilde, selv når de er belagt med Poly-L-lysine. Videre, de er mer sårbar enn somatiske celler og vev inndelinger som resulterer i cellen lysis når utsatt for noen av proprietære bufferne i kommersielt tilgjengelig kits3. For å overvinne disse utfordringene, oocytes fast og manuelt overføres mellom dråper bufferne. Videre erstattet permeabilization og vask buffere i settene for å redusere celle lysis. Forhåndsutformede sonder kjøpes sammen med fisk kit eller bestemte utskrifter kan forespørres. Hver proprietære sonde er tilgjengelig i en av tre fluorescens kanaler (C1, C2 og C3) for å tillate multipleksing. I gjeldende eksperimentet var murine oocytes dual-farget og kvantifisert ved hjelp av en C2 Nanog sonde og en C3 Pou5f1 sonde. Disse sonder ble valgt basert på rapporterte uttrykk for Nanog og Pou5f1 i oocytes og embryo. Ved avslutningen av hybridisering trinnene plassert oocytes i dråper av anti-fade monterer medier skal histologiske lysbilder. AC confocal bilder ble brukt om å kvantifisere antall vises punctate fluorescerende signaler som representerer personlige mRNAs. Kvantifisere mRNAs, bildebehandling viste også den romlige fordelingen av den spesifikke mRNA i cellen, er hvilke andre RNA kvantifisering metoder ikke å oppnå. Denne teknikken vist seg for å ha lav variasjon innen en forsøksgruppen tillater bruk av mindre antall oocytes i hver forsøksgruppen å identifisere betydelige forskjeller mellom eksperimentelle grupper3.
