Substratos estáticos e dinâmicos são duas categorias de biomateriais que foram desenvolvidas para estudar os efeitos de elasticidade do tecido, ou em função da rigidez das células. Substratos estáticos são incapazes de alterar as suas propriedades físicas depois de serem fabricadas e / ou uma vez as células são banhados. Poliacrilamida (PA) foram os primeiros géis bidimensionais substratos, estáticas que foram sintetizadas para investigações mechanobiology 5,17. Geles PA são fáceis de preparar, barato, versátil e pode ser fabricado com uma ampla gama de módulos elásticos. Embora essas vantagens técnicas fazem PA géis um substrato comumente aplicado, substratos estáticas não são indicativos da natureza dinâmica da matriz extracelular (MEC) e ao ambiente circundante celular in vivo. Por exemplo, a rigidez ECM sofre alterações em resultado de ferimento, de desenvolvimento, ou doença. Substratos dinâmicos são, portanto, preferidas como substratos modelo mimetizando a tecidos em estudos mechanobiology <sup> 22,24,25.
Numerosos sintético, Bidimensional biomateriais, tridimensionais, estáticos e dinâmicos naturais têm sido desenvolvidos para imitar rigidez tecidual 1,3,6,16,23,26. Alguns substratos dinâmicas exigem calor, UV, corrente elétrica, os íons, e mudanças de pH de alterar as suas propriedades mecânicas 2,4,7,8,12,15,16, mas esses estímulos podem restringir bio-aplicação do hidrogel. Hidrogeles de poliacrilamida reticulada-DNA (DNA) são géis elásticos substratos bidimensionais dinâmicas. Ligações cruzadas de DNA permitem a modulação temporais, espaciais, e reversível da rigidez gel DNA pela adição de DNA de fita simples (ssDNA) para mídia ou buffer 9-11,13,14,18,21. Ao contrário dos géis dinâmicas acima mencionados, onde são aplicados estímulos para modulação de elasticidade, os géis de DNA contam com a difusão de ADNcs aplicada para a alteração da elasticidade. Por conseguinte, a superfície superior de gel, em que as células são cultivadas, é a primeira área modulada, porque a taxa de omodulação f elasticidade está dependente da espessura do gel.
Geles de DNA são semelhantes aos seus homólogos gel PA na medida em que têm uma espinha dorsal de poliacrilamida, no entanto, as ligações cruzadas de bis-acrilamida substituída com ligações cruzadas são compostos por ADN (Figura 1). Dois ssDNAs (SA1 e SA2) hibridar com uma cadeia reticulador (L2) para compensar as ligações cruzadas de DNA do gel. SA1 e SA2 têm seqüências distintas que ambos contêm uma modificação Acrydite no final 5 'para a incorporação efetiva na rede PA. Para a preparação do gel, SA1 e SA2 são polimerizados, individualmente, em uma espinha dorsal PA e, subsequentemente, o SA1 polimerizada e SA2 são misturados em conjunto. L2, o agente de reticulação, é adicionado à mistura de SA1 e SA2. A sequência de bases é complementar a L2 ambas as sequências SA1 e SA2 e L2 hibrida com SA1 mais SA2 para formar as ligações cruzadas de DNA. Inicial, a elasticidade de gel de ADN é determinada por ambas as concentrações de L2 e de reticulação (Tabelas 1 </strong> e 2). Geles de ADN que contenham quantidades estequiométricas iguais de L2, AE1, AE2 e os géis são mais duras, porque SA1 e SA2 são 100% reticulado por L2 (designado como 100% de gel). Baixas concentrações do resultado L2 em uma menor percentagem de reticulação DNA e, portanto, géis de DNA mais suave. Géis tão baixas como 50% reticulado (designado como 50% de gel) 9-11 foram construídas.
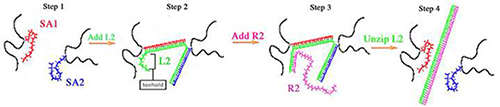
A Figura 1 de reticulação de gel do DNA e uncrosslinking esquemática 9-11,13,14,18,21 Etapa 1:. SA1 (vermelho) e SA2 (azul) são polimerizados, individualmente, em uma espinha dorsal de poliacrilamida (preto). Após a polimerização, as soluções polimerizados SA1 e SA2 são misturados em conjunto. Passo 2: L2 (verde) é adicionado e hibrida com SA1 mais SA2 para formar as ligações cruzadas do gel. Passo 3: R2 hibrida com tele toehold de L2. Passo 4: hibridização toehold de R2 impulsiona a descompactação de L2 de SA1 e SA2.
Ao contrário dos géis PA, géis de DNA pode endurecer e amaciar após a síntese. Por essa razão, as células cultivadas em géis de ADN pode ser submetido a alterações de rigidez dinâmica. Para endurecer géis de células aderentes, L2 pode ser adicionado ao meio de cultura de baixo géis percentuais, para aumentar a percentagem de ligações cruzadas. Para suavizar a géis de células aderentes, L2 podem ser removidas para reduzir a percentagem de ligações cruzadas 10,13,21. L2 tem uma sequência de ponto de apoio adicional na extremidade 3 'para permitir a L2 uncrosslink de SA1 e SA2 (Tabela 1). Remoção de L2 é realizado através de hibridação de um fio chamado inversão R2. R2 é complementar ao comprimento total de L2 e hibrida com o primeiro ponto de apoio L2. Toehold hibridação impulsiona a descompactação de L2 de SA1 e SA2, que elimina a ligação transversal e reduz a rigidez do gel.
Neste relatório evídeo, passo-a-passo as instruções são fornecidas para a preparação de endurecimento e amolecimento géis de DNA. Embora 100% e de 80% as preparações de gel são descritas, este protocolo pode ser adaptada para criar ADN de géis reticulados outras percentagens iniciais e finais. Em geral, 100% e 80% de gel são preparadas, imobilizada em lamelas de vidro, funcionalizada, e semeadas com células. L2 é adicionado à media de 80% de gel e R2 é adicionado aos meios de comunicação de 100% de gel, 48 horas após o plaqueamento. A adição de meios de L2 para endurece 80% de gel de ligação cruzada a 100%, ao passo que a adição de R2 a mídia amolece a 100% de gel de 80% de ligações cruzadas. Geles endurecidos são designados como 80 → 100% de gel e geles suavizados são designados como 100 → 80% de gel no texto. Para o controlo ou géis estáticos, ADNcs ou consistindo em Ts Como é entregue a outro conjunto de 100% e 80% de gel. Depois de um mínimo de dois dias seguintes modulação elasticidade, as células podem ser processados e analisados.
<table border="0" cellpadding="0" cellspacing="0" fo:keep-together.within-page = "always">Tabela 1. seqüências de bases para ssDNA 9-11,13,14,18,21. Celular e estudos mecânicos têm utilizado vários dmodelos de reticulação iferentes para gerar géis de ADN com uma variedade de propriedades mecânicas estáticas e dinâmicas. Os parâmetros modulados em design de reticulação são sequência de bases de comprimento e sequência ou comprimento de reticulação. Fontes em negrito e itálico ilustrar o pareamento de bases entre SA1 e L2 e entre SA2 e L2, respectivamente.
| Projeto | ||||||||
| 1 | 2 | 3 | ||||||
| Concentração de acrilamida (%) | 10 | 10 | 10 | 4 | ||||
| SA1 mais SA2 hibridado com L2 (% reticulado) | 50 | 80 | 100 | 50 | 80 | 100 | 100 | 100 |
| <strong> Elasticidade (kPa, média ± SEM) | 6,6 ± 0,6 | 17,1 ± 0,8 | 29,8 ± 2,5 | 5,85 ± 0,62 | 12,67 ± 1,33 | 22,88 ± 2,77 | 25,2 ± 0,5 | 10,4 ± 0,6 |
Tabela 2 o módulo de Young (E) de geles de ADN. 9-11,13,14,18,21 concentração de acrilamida, percentagem de ligações cruzadas, e comprimento de ligação transversal pode ser modulada em geles de ADN. Designs 1, 2 e 3 têm 20, 28 e 40 pb comprimentos de ligação transversal, respectivamente. 100% de gel para todos os projetos têm módulos semelhantes indicando comprimento reticulação não afeta a elasticidade gel. No entanto, as variações na concentração de acrilamida alterar a elasticidade em gel de ADN.
