Sustratos estáticos y dinámicos son dos categorías de biomateriales que se desarrollaron para estudiar los efectos de la elasticidad de los tejidos o rigidez en la función celular. Sustratos estáticas son incapaces de cambiar sus propiedades físicas después de que se fabrican y / o una vez se siembran las células. Poliacrilamida (PA) geles fueron los primeros sustratos bidimensionales estáticas, que se sintetizaron las investigaciones mecanobiología 5,17. Geles de PA son fáciles de preparar, de bajo costo, versátil, y puede ser fabricado con una amplia gama de módulos elásticos. Aunque estas ventajas técnicas hacen PA geles un sustrato general aplicada, sustratos estáticas no son indicativos de la naturaleza dinámica de la matriz extracelular (MEC) y que rodea entorno celular in vivo. Por ejemplo, el ECM sufre alteraciones de rigidez como resultado de una lesión, el desarrollo, o la enfermedad. Por lo tanto sustratos dinámicos son favorecidos como los modelos de sustrato que imitan los tejidos en los estudios mecanobiología <sup> 22,24,25.
Numerosos sintética, bidimensional biomateriales, tridimensionales, estáticas y dinámicas naturales se han desarrollado para imitar la rigidez del tejido 1,3,6,16,23,26. Algunos sustratos dinámicos requieren calor, UV, la corriente eléctrica, los iones, y los cambios de pH para alterar sus propiedades mecánicas 2,4,7,8,12,15,16, pero estos estímulos pueden restringir bio-aplicación del hidrogel. Hidrogeles de poliacrilamida reticulada de ADN-ADN (geles) son sustratos elásticos bidimensionales dinámicos. Entrecruzamientos de ADN permiten la modulación temporal, espacial, y reversible de la rigidez en gel de ADN mediante la adición de ADN de cadena sencilla (ssDNA) a los medios o tampón 9-11,13,14,18,21. A diferencia de los geles dinámicas mencionadas anteriormente donde se aplican los estímulos para la modulación de elasticidad, los geles de ADN se basan en la difusión de ssDNA aplicada para la alteración de la elasticidad. Por lo tanto, la superficie del gel superior, donde se cultivan las células, es la primera área modulada porque el o tasamodulación f elasticidad es dependiente del espesor de gel.
Geles de ADN son similares a sus homólogos de gel de PA en que tienen un esqueleto de poliacrilamida, sin embargo, las reticulaciones-bis acrilamida se reemplazan con reticulaciones compuestas de ADN (Figura 1). Dos ssDNAs (SA1 y SA2) se hibridan con una cadena reticulante (L2) para compensar los entrecruzamientos de ADN del gel. SA1 y SA2 tienen secuencias distintas que ambos contienen una modificación Acrydite en el extremo 5 'para su incorporación efectiva a la red PA. Para la preparación de los geles, SA1 y SA2 se polimerizan individualmente en una cadena principal de PA y, posteriormente, el SA1 SA2 polimerizado y se mezclan juntos. L2, el agente de reticulación, se añade a la SA1 y SA2 mezcla. La secuencia de bases es complementaria a L2 ambas secuencias SA1 y SA2 y L2 se hibrida con SA1 SA2 más para formar los entrecruzamientos de ADN. , Elasticidad del gel de ADN inicial es determinado por ambas concentraciones L2 y reticulación (Tablas 1 </strong> y 2). Geles de ADN que contienen cantidades estequiométricas iguales de L2, SA1, SA2 y son los geles más rígidas porque SA1 y SA2 son 100% reticulado por L2 (designado como 100% geles). Concentraciones más bajas de L2 resultado en un menor porcentaje de reticulación del ADN y, por tanto, geles más blandos de ADN. Geles tan bajas como 50% reticulado (designado como 50% en geles) se han construido 9-11.
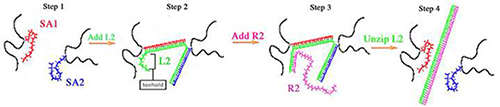
Figura 1. reticulación de gel de ADN y uncrosslinking esquemática 9-11,13,14,18,21 Paso 1:. SA1 (rojo) y SA2 (azul) se polimerizan de forma individual en una columna vertebral de poliacrilamida (negro). Después de la polimerización, soluciones polimerizadas SA1 y SA2 se mezclan juntos. Paso 2: L2 (verde) se añade y se hibrida con SA1 SA2 más para formar los entrecruzamientos del gel. Paso 3: R2 hibrida con tél punto de apoyo de la L2. Paso 4: hibridación de punto de apoyo de R2 impulsa la descompresión de la L2 de SA1 y SA2.
A diferencia de los geles de PA, geles de ADN pueden endurecerse y suavizar después de la síntesis. Por esa razón, las células cultivadas en geles de ADN pueden ser sometidos a alteraciones de rigidez dinámica. Para endurecer geles de células adherente, L2 puede ser añadido al medio de cultivo de los geles porcentuales bajos para aumentar el porcentaje de reticulaciones. Para suavizar los geles de células adherente, L2 se puede quitar para disminuir el porcentaje de entrecruzamientos 10,13,21. L2 tiene una secuencia de punto de apoyo adicional en el extremo 3 'para permitir L2 para uncrosslink de SA1 y SA2 (Tabla 1). La eliminación de L2 se lleva a cabo mediante hibridación de una hebra de reversión llamado R2. R2 es complementaria a la longitud completa de L2 y se hibrida primero con el punto de apoyo L2. Hibridación propulsa el punto de apoyo de descompresión de L2 de SA1 y SA2, que elimina la reticulación y reduce la rigidez de gel.
En el presente informe yvideo, paso a paso las instrucciones se proporcionan para la preparación de rigidización y suavizar los geles de ADN. Si bien se describen preparaciones de gel 100% y 80%, este protocolo se puede adaptar para crear geles de ADN de otros porcentajes reticulados inicial y final. En general, 100% y 80% en geles se preparan, inmovilizan sobre cubreobjetos de vidrio, funcionalizados, y se sembraron con células. L2 se añade a los medios de 80% de geles y R2 se añade a los medios de comunicación de 100% geles, 48 hr después de la siembra. La adición de L2 a los medios endurece 80% a 100% geles reticulados, mientras que la adición de R2 a los medios suaviza 100% en geles de 80% reticulado. Geles endurecidas se designan como 80 → 100% geles y geles ablandados se designan como 100 → 80% de geles en el texto. Para controlar o geles estáticas, ssDNA que consiste en Ts o Como se entrega a otro conjunto de 100% y 80% en geles. Después de un mínimo de dos días siguientes modulación de elasticidad, las células pueden ser procesados y analizados.
<table border="0" cellpadding="0" cellspacing="0" fo:keep-together.within-page = "always">Cuadro 1 secuencias de base para ssDNA 9-11,13,14,18,21. Celular y estudios mecánicos han utilizados varios ddiseños de reticulación ifferent para generar geles de ADN con una gama de propiedades mecánicas estáticas y dinámicas. Los parámetros modulados en el diseño de reticulación son secuencia de bases y longitud de la secuencia o la longitud de reticulación. Negrita y cursiva ilustran base de sincronización entre SA1 y L2 y entre SA2 y L2, respectivamente.
| Diseño | ||||||||
| 1 | 2 | 3 | ||||||
| Concentración de acrilamida (%) | 10 | 10 | 10 | 4 | ||||
| SA1 SA2 más hibridado con L2 (% reticulado) | 50 | 80 | 100 | 50 | 80 | 100 | 100 | 100 |
| <strong> Elasticidad (kPa, media ± SEM) | 6,6 ± 0,6 | 17,1 ± 0,8 | 29,8 ± 2,5 | 5,85 ± 0,62 | 12.67 ± 1.33 | 22.88 ± 2.77 | 25,2 ± 0,5 | 10,4 ± 0,6 |
Tabla 2. módulo de Young (E) de geles de ADN. 9-11,13,14,18,21 concentración de acrilamida, el porcentaje de reticulación, y la longitud de reticulación puede ser modulada en geles de ADN. Diseños 1, 2 y 3 tienen 20, 28, y 40 pb longitudes de reticulación, respectivamente. 100% de geles para todos los diseños tienen módulos similares, indicando la longitud de reticulación no afecta a la elasticidad del gel. Sin embargo, las variaciones en la concentración de acrilamida alteran elasticidad del gel del ADN.
